Chromatin and its types
Chemically chromatin consists of DNA, RNA, protein and inorganic ions Mg2+, Ca2+. The protein of chromatin could be of two types: histones and non-histones.
DNA
DNA is the most important chemical component of chromatin since it plays the central role of controlling heredity and is most conveniently measured in picograms. In addition to describing the genome of an organism by its number of chromosomes, it is also described by the amount of DNA in a haploid cell. Lower eukaryotes, in general, have less DNA, such as nematode Caenorhabditiselegans which has 20 times more DNA than E. coli. Vertebrates have greater DNA content about 3pg, in general about 700 times more than E. coli. Salamander amphiuma has a very high DNA content of about 84pg. Man has about 3pg of DNA per haploid genome.
Histones
Histones are basic proteins as they are enriched with basic proteins arginine and lysine. At physiological pH, they are cationic and can interact with anionic nucleic acids. They form a highly condensed structure. The histones are of five types called H1, H2A H2B, H3, and H4 which are very similar among different species of eukaryotes and have been highly conserved during evolution. H1 is the least conserved among all and is also loosely bound with DNA. H1 histone is absent in Saccharomyces cerevisiae. All histones can be separated by gel electrophoresis. Histones are basic proteins as these contain a high proportion of basic amino acids.
Non-histones
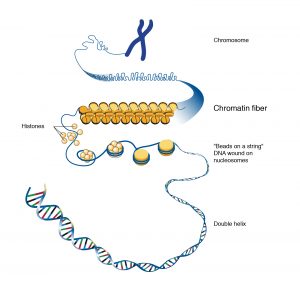
In addition to histones, the chromatin comprises of many different types of nonhistone proteins, which are involved in a range of activities, including DNA replication and gene expression. They display more diversity or are not conserved. They may also differ between different tissues of the same organism. Non-histone proteins associated with DNA are acidic in nature and include various enzymes and proteins involved in the process of replication and transcription. Roger Kornberg in 1974 described the basic structural unit of chromatin which is called the nucleosome. DNA is folded into bead-like structures called nucleosomes, which are repeating units of chromosome structure. These units are remarkably uniform in size, about 11 nm in diameter.
In general terms, there are three levels of chromatin organization:
- DNA wraps around histone proteins forming nucleosomes; the “beads on a string” structure (euchromatin).
- Multiple histones wrap into a 30 nm fibre consisting of nucleosome arrays in their most compact form (heterochromatin).
- Higher-level DNA packaging of the 30 nm fibre into the metaphase chromosome (during mitosis and meiosis).
Types of chromatin
Two types of chromatin are found: 1.Euchromatin and 2. Heterochromation
- Euchromatin: The lightly-stained regions in chromosome when stained with basic dyes are called euchromatin and contain single-copy of genetically-active DNA. The extent of chromatin condensation varies during the life cycle of the cell and plays an important role in regulating gene expression. It is transcriptionally active chromatin. In the interphase of the cell cycle, the chromatin is decondensed and known as euchromatin leading to gene transcription and DNA replication.
- Heterochromatin: The word heterochromatin was coined by Emil Heitz based on cytological observations. They are highly condensed and ordered areas in nucleosomal arrays. About 10% of interphase chromatin is called heterochromatin and is in a very highly condensed state that resembles the chromatin of cells undergoing mitosis. They contain a high density of repetitive DNA found at centromeres and telomeres from heterochromatin. Heterochromatin is of two types, the constitutive and facultative heterochromatin. The regions that remain condensed throughout the cell cycle are called constitutive heterochromatin whereas the regions where heterochromatin condensation state can change are known as facultative. Heterochromatin is transcriptionally inactive and late replicating or heteropycnotic. Constitutive heterochromatin is found in the region that flanks the telomeres and centromere of each chromosome and in the distal arm of the Y chromosome in mammals. Constitutive heterochromatin possesses very few genes and they also lead to transcriptional inactivation of nearby genes. This phenomenon of gene silencing is known as “position effect”. Constitutive heterochromatin also inhibits genetic recombination between homologous repetitive sequences circumventing DNA duplications and deletion. Whereas facultative heterochromatin is the chromatin that has been specifically inactivated during certain phases of an organism’s life or in certain types of differentiated cells. Dosage compensation of X-chromosome or X-chromosome inactivation in mammals is an example of such heterochromatin (Karp 2010). Heterochromatin spreads from a specific nucleation site, causing silencing of most of the X chromosome, thereby regulating gene dosage.