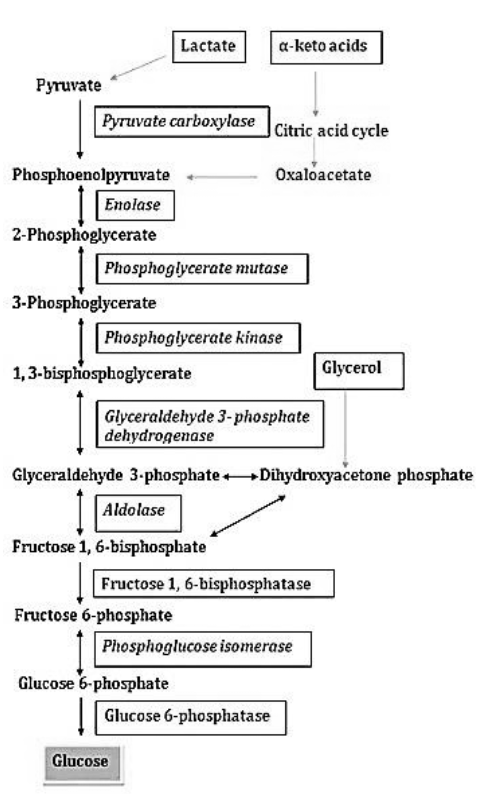
Gluconeogenesis Pathway
Some tissues, such as the brain and erythrocytes, depend on a constant supply of glucose. If the amount of carbohydrate taken up in food is not sufficient, the blood sugar level can be maintained by degradation of glycogen (a polymer of glucose) for a limited time. If these reserves are also exhausted, the de-novo synthesis of glucose begins, which is known as gluconeogenesis. The major precursors for gluconeogenesis are lactate, pyruvate, glycerol, and some amino acids.
Location of pathway
Gluconeogenesis occurs mainly in cytosol, although some precursors are produced in mitochondria. It occurs in all animals, plants, fungi, and microorganisms. In mammals, gluconeogenesis takes place mainly in the liver and to a lesser extent in the matrix of the kidney.
Reactions of pathway
Gluconeogenesis and glycolysis are not identical pathways running in opposite directions, although they do share several steps; seven of the ten enzymatic reactions of gluconeogenesis are the reverse of glycolytic reactions. However, three reactions of glycolysis are essentially irreversible and cannot be used in gluconeogenesis are:
a) The conversion of glucose to glucose 6-phosphate by hexokinase
b) The phosphorylation of fructose 6-phosphate to fructose 1,6-bisphosphate by PFK
c) The conversion of phosphoenolpyruvate to pyruvate by pyruvate kinase
These three steps of glycolysis are bypassed in gluconeogenesis which is catalyzed by some specific enzymes.
a) Conversion of pyruvate to phosphoenolpyruvate
- This conversion is taking place in two steps. Pyruvate carboxylase, a mitochondrial enzyme converts pyruvate to oxaloacetate in presence of ATP and CO2. This is the regulatory enzyme of gluconeogenesis which requires acetyl CoA for its activity.
- Oxaloacetate is synthesized in the mitochondrial matrix. It has to be transported to the cytosol to be used in gluconeogenesis, where the rest of the pathway occurs.
- Due to membrane impermeability, oxaloacetate cannot diffuse out of the mitochondria.
- It is converted to the malate and then transported to the cytosol. Within the cytosol, oxaloacetate is regenerated. The reversible conversion of oxaloacetate and malate is catalyzed by malate dehydrogenase.
- In the cytosol, phosphoenolpyruvate carboxykinase converts oxaloacetate to phosphoenolpyruvate, GTP is used in this reaction and CO2 is liberated.
- The subsequent steps up to fructose 1,6-bisphosphate represent the reverse of the corresponding reactions involved in glycolysis.
b) Conversion of fructose 1,6-bisphosphate to fructose 6-phosphate
The enzyme fructose 1,6-bisphosphatase converts fructose 1,6-bisphosphate to fructose 6-phosphate. This enzyme requires Mg” for its activity. It is also one of the regulatory enzymes in gluconeogenesis.
c) Conversion of glucose 6-phosphate to glucose
- The hexokinase reaction of glycolysis is irreversible, the hydrolysis of glucose 6-phosphate to glucose is carried out by glucose 6-phosphatase in the final step of the pathway.
- The presence or absence of this enzyme in a tissue determines whether the tissue is capable of contributing glucose to the blood or not. It is mostly present in the liver and kidney but absent in muscle, brain, and adipose tissue.
- Overall summary of gluconeogenesis
2 Pyruvate +4ATP +2GTP+2NADH +2H+ +6H2O ——> Glucose +2NAD+ +4ADP +2GDP + 6Pi+ 6H+
- For each molecule of glucose formed from pyruvate, six high-energy phosphate groups are required, four from ATP and two from GTP. In addition, two molecules of NADH are required for the reduction of two molecules of 1,3-bisphosphoglycerate.
- The synthesis of glucose from pyruvate is a relatively expensive process. Much of this high energy cost is necessary to ensure the irreversibility of gluconeogenesis.