Plasma Membrane is the boundary that separates the living cell from its nonliving surroundings. All cells-both prokaryotic and eukaryotic -are surrounded by a plasma membrane, which defines the boundary of the cell. The plasma membrane exhibits selective permeability, allowing some substances to cross easily while it is difficult for certain substances. It is around ~8 nm thick. It is impermeable to most water-soluble molecules. The passage of ions and most biological molecules across the plasma membrane is therefore mediated by proteins, which are responsible for the selective traffic of molecules into and out of the cell. Other proteins of the plasma membrane control the interactions between cells of multicellular organisms and serve as sensors through which the cell receives signals from its environment.
Structure of the Plasma Membrane
- In 1935, Hugh Davson and James Danielli proposed a sandwich model in which the phospholipid bilayer lies between two monolayers of globular proteins.
- In 1965 Robertson proposed the Unit membrane hypothesis which was later found to have any problems with the passage of molecules and placement of membrane proteins and, which have hydrophilic and hydrophobic regions.
- In 1972, S. J. Singer and G. Nicolson proposed the fluid mosaic model which suggests that the membrane is a mosaic of proteins dispersed within the bilayer, with only the hydrophilic regions exposed to water. It is the most accepted model for the plasma membrane.

Components of the cell membrane
- Lipid→ cholesterol, phospholipid, and sphingolipid
- Proteins
- Carbohydrate – as glycoprotein Differences in composition among membranes (e.g. myelin vs. inner mitochondrial membrane) This is due to the differences in function. Example: Mitochondrial inner membrane has high amounts of functional electron transport system proteins.
- variability of membrane structure.
The plasma membrane, with fewer functions (mainly ion transport), has less protein.
- Membranes with similar functions (i.e. from the same organelle) are similar across species lines, but membranes with different functions (i.e. from different organelles) may differ strikingly within a species.
Carbohydrates of membranes are presently attached to protein or lipid as glycoprotein or glycolipid.
- Typical sugars in glycoproteins and glycolipids include glucose, galactose, mannose, fucose, and N-acetylated sugars like N-acetylglucosamine, N-acetylgalactosamine, and N-acetylneuraminic acid (sialic acid).
- Membrane sugars seem to be involved in identification and recognition.
The Phospholipid Bilayer
The plasma membranes of mammalian red blood cells (erythrocytes) have been particularly useful as a model for studies of membrane structure. Mammalian red blood cells do not contain nuclei that serve as pure plasma membranes that can be easily isolated for biochemical analysis.
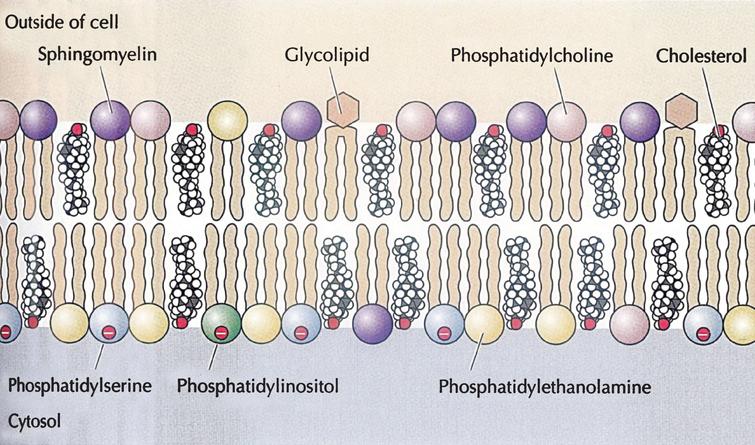
Phospholipids are the most abundant lipid in the plasma membrane (sphingolipid, glycolipid, and steroid). Phospholipids are amphipathic molecules, containing hydrophobic and hydrophilic regions. The plasma membranes of the animal cell contain (phosphatidylcholine, phosphatidylethanolamine, phosphatidylserine, and sphingomyelin).
The amphipathic properties of the phosphoglycerides and sphingolipids are due to their structures.
- The hydrophilic head bears electric charges contributed by the phosphate and by some of the bases.
- These charges are responsible for hydrophilicity.
- Note that no lipid bears a positive charge. They are all negative or neutral. Thus membranes are negatively charged.
- The long hydrocarbon chains of the acyl groups are hydrophobic and tend to exclude water.
- Phospholipids in an aqueous medium spontaneously aggregate into orderly arrays.
- Micelles: orderly arrays of molecular dimensions. Note the hydrophilic heads oriented outward, and the hydrophobic acyl groups oriented inward. Micelles are important in lipid digestion; in the intestine, they assist the body in assimilating lipids
- Lipid bilayers can also form.
- Liposomes are structures related to micelles, but they are bilayers, with an internal compartment. Thus there are three regions associated with liposomes: -The exterior, the membrane itself, and the inside.
- Liposomes can be made with specific substances dissolved in the interior compartment. These may serve as modes of delivery of these substances.
- The properties of phospholipids determine the kinds of movement they can undergo in a bilayer.
- Modes of movement that maintain the hydrophilic head in contact with the aqueous surroundings and the acyl groups in the interior are permitted.
- Transverse movement from side to side of the bilayer (flip-flop) is relatively slow and is not considered to occur significantly.
The outer leaflet of the plasma membrane consists mainly of phosphatidylcholine and sphingomyelin, whereas phosphatidylethanolamine and phosphatidylserine are the predominant phospholipids of the inner leaflet which have negative head groups making it negatively charged.
A fifth phospholipid, phosphatidylinositol, is also localized to the inner half of the plasma membrane which has a major role in cell signaling. In addition to the phospholipids, the plasma membranes of animal cells contain glycolipids (2%) and cholesterol
General Features Of Phospholipids
- Phospholipids are responsible for the function of membranes to act as barriers between two aqueous compartments. The inner leaflet of phospholipid bilayer is hydrophobic fatty acid chains that are impermeable to water-soluble molecules, ions, and most biological molecules.
- The phospholipids are vicious in nature which allows free movement of hydrocarbon chains making in soft and flexible. Both phospholipids and proteins are free to diffuse laterally within the membrane.
Cholesterol
Cholesterol plays a distinct role in membrane structure. It inserts into a bilayer of phospholipids with its polar hydroxyl group close to the phospholipid head groups. The temperature controls cholesterol activity. At high temperatures, it interferes with the movement of the outer membrane restricting permeability to small molecules whereas at low temperatures it interferes with interactions between fatty acid chains and prevents membranes from freezing with maintaining membrane fluidity. Plant cell maintains the cholesterol function by sterols which act the same way.
Rather than diffusing freely in the plasma membrane, cholesterol and the sphingolipids (sphingomyelin and glycolipids) form discrete membrane domains known as lipid rafts. These clusters of sphingolipids and cholesterol move laterally within the plasma membrane and associate with specific membrane proteins. Although the functions of lipid rafts remain to be fully understood, they play important roles in processes such as cell movement and the uptake of extracellular molecules by endocytosis as well as in cell signaling.
Membrane Proteins and Their Functions
Membrane proteins are responsible for major membrane functions.
After the proposal of the fluid mosaic model by Singer and Nicolson, they explained two major membrane proteins.
- Peripheral Proteins
- Integral Proteins
- Peripheral proteins are bound to the surface of the membrane
- Integral proteins that penetrate the hydrophobic core that spans the membrane are called transmembrane proteins. The hydrophobic regions of integral protein consist of one or more stretches of nonpolar amino acids, often coiled into alpha-helices
Example: Band 3 Protein
An example of an intrinsic membrane protein is Band 3 protein in the human plasma cell membrane. Band 3 has a composition of two domains. In the cytosol, the amino-end of 41 kDa is situated and the cytoskeleton is binding to the membrane. The 55 kDa domain carboxy-terminal is integrated into the lipid bilayer and is responsible for the transportation of anion. RBC membrane is a two-dimensional structure consisting basically of a cytoskeleton and a lipid bilayer, tied together. In the spectrin ankyrin binding sites and via glycophorin, the cytoskeleton is tied to the lipid bilayer through immobile band-3 protein at the actin junctional complexes.
Functions
The six major functions of membrane proteins are Transport, Enzymatic activity, Signal transduction, Cell-cell recognition, Intercellular joining, Attachment to the cytoskeleton, and extracellular matrix (ECM).
Mobility of Membrane Proteins
Membrane proteins and phospholipids are unable to move back and forth between the inner and outer leaflets of the membrane at an appreciable rate but they can diffuse laterally through the membrane.
However, not all proteins can diffuse freely through the membrane. In some cases, the mobility of membrane proteins is restricted by their association with the cytoskeleton. For example, a fraction of band 3 in the red blood cell membrane is immobilized as a result of its association with ankyrin and spectrin.
References
- https://www.ncbi.nlm.nih.gov/books/NBK9898/
- https://www.ncbi.nlm.nih.gov/books/NBK26871/
- https://www.ncbi.nlm.nih.gov/pmc/articles/PMC4129483/
- Cell and Molecular Biology by Karp 5th Ed
- Karp’s Cell and Molecular Biology: Concepts and Experiments, 5th Edition Gerald Karp, Janet Iwasa, Wallace Marshall