Temperature relationship Definitions
Animals are limited to fairly narrow temperature ranges (approximately –2°C to +50°C for internal temperatures which will sustain active metabolism). Most animals (including all aquatic invertebrates) have the same temperatures as their surroundings. Birds and mammals keep their body temperatures approximately constant, and many other animals maintain a substantial difference between their own body (or parts of their body) temperature and that of the environment.
Definitions
● Warm-blooded (e.g. mammal) – but a hibernating mammal may be at 4°C.
● Cold-blooded (e.g. lizard) – but a lizard basking may be at more than 40°C.
● Poikilothermic (= changeable temperature) – but deep-sea fish may be at a very constant temperature throughout life.
● Homoiothermic (= constant temperature) – but this may change in a hibernating hedgehog: the term heterothermic may be preferable for hibernating homoiotherms.
● Ectothermic – relies on external sources of heat (e.g. lizard, but neonatal mice may be temporary ectotherms).
● Endothermic – relies on internally generated, metabolic heat (e.g. bird, but shark and tuna muscles and moth flight muscles may conserve metabolic heat).
Thus these definitions obviously have their limitations
Effects of temperature change
Within limits, raised temperatures to accelerate most physiological processes: oxygen consumption increases as the metabolic rate of the animal increases. Within physiological limits, a rise of 10°C leads to an increase in oxygen consumption (and therefore of metabolic rate) by a factor of 2–3. The increase in rate over 10°C = Q10. Therefore, if Q10 = 2, the rate doubles over 10°C. Thus, if the rate = x at 0°C, it will be 2x at 10°C, it will be 4x at 20°C, it will be 8x at 30°C and so on; that is, there is an exponential increase. In practice, Q10 values are usually between 2 and 3; for example for a metabolic rate in a Colorado beetle between 7 and 30°C, Q10 = 2.17. (The Q10 value falls at higher temperatures, particularly as enzyme denaturation starts to occur.) By contrast, the Q10 values for physicochemical reactions are generally less than 1.2.
Temperature extremes
Some animals can tolerate only a narrow range of temperatures (stenothermic), others a wide range (eurythermic). Tolerances can change with time (with less tolerance when immature). There is a distinction between temperatures at which an animal can survive and those at which it can carry out its entire life cycle. The lethal temperature is often difficult to define and exposure time can be important, as for humans in a sauna bath.
High temperature tolerance
There is no known animal which can complete its life cycle at more than 50°C, but resting stages can be very tolerant: the larva of the fly Polypedium sp. can dehydrate and survive 102°C for 1 minute. Triops sp. eggs (a Sudanese crustacean) can estivate in mud during drought at 80°C and will survive short periods at even higher temperatures in the laboratory. Humidity is important: cockroaches will survive for 24 hours at 37°C in damp air but will die at the same temperature in dry air.
Many animals die when exposed to temperatures above 50°C. Marine forms would be exposed to temperatures greater than 50°C only if they were in the intertidal zone. For snails, heat resistance reflects their position in the intertidal zone. The freshwater desert pupfish from Devil’s Hole, California lives in water at a constant 34°C – its upper lethal limit is 43°C; the antarctic ice-fish Trematomus sp., living in water at –1.9 ± 0.1°C, has an upper lethal limit of 6°C. Heat death is probably due to different Q10 values for interdependent metabolic reactions: changes in temperatures unbalance the ordered sequence of reactions and the proportions and availability of (intermediate) products. Extreme high temperatures lead to enzyme denaturation.
Low-temperature tolerance
Some animals will tolerate freezing, but the lower lethal limits for others is higher: a guppy kept at 23°C dies at 10°C – the respiratory centres of the hindbrain become anaesthetized, leading to anoxia.
Cold and freezing tolerance may be due to:
● Supercooling (cooling below 0°C at atmospheric pressure without freezing): water can be supercooled, dependent on the temperature, time and the absence of nuclei for ice crystal formation. Solutes may enhance supercooling. It may be important for a lizard exposed to –6°C when caught out on a cold night.
● Freezing tolerance: a few animals can tolerate ice crystals (and the resulting high concentrations of salts). Chironomus larvae will survive repeated freezing and thawing to –25°C. It may be important for intertidal organisms in winter polar seas. Ice crystals are outside of the cells which are distorted. Freezing tolerance varies with season.
● Antifreezes: glycerol is found in some insect hemolymphs. It may protect cells against ice crystal damage, lower freezing points and promote supercooling.
In the wasp Bracon cephi, glycerol concentrations rise to 30% in the autumn, giving a freezing point of –17.5°C; supercooling permits survival below –47°C. In teleost fishes, supercooling is noted and there is evidence for antifreezes. In Trematomus sp. the antifreeze is a glycoprotein.
Tolerance limits will vary according to acclimatization (artificially induced acclimatization = acclimation), season, age, geographical distribution, etc. Arctic temperatures are lethal to tropical species as are tropical temperatures to polar species.
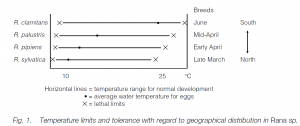
Even closely related species (e.g. Rana spp.) have tolerances related to their geographical distributions. The teleost bullhead has an upper lethal limit of 28°C in winter and 36°C in summer. If bullheads are acclimated to a changed temperature, the shift in temperature tolerance is rapid when the temperature of acclimation is moved upwards, but slower when it is moved downwards.