Cleavage :
After fertilization the activated zygote, cell passes through a series of mitotic cell division in a rapid, succession, so that unicellular body is transformed into a complex, multicellular body of the offspring. This transformation, take place in the beginning of development and is called as cleavage or segmentation.
Characteristics:
- The fertilized egg divides repeatedly by mitosis to produce thousands of cells called blastomeres.
- Cleavage produces a solid mass of cells called morula. Morula develops only in certain animals. Eg. Mammals.
- In majority of animals cleavage produces a hollow sphere of cells called blastula.
- Cleavage provides an adequate number of cells for the construction of tissues and organs.
- During cleavage, the embryo does not grow.
- The shape of the embryo does not change during cleavage.
- During cleavage the amount of DNA increases.
- Cleavage brings about a fixed proportion between nuclear and cytoplasmic materials.
Significance:
The significance of cleavage comprises the following aspects. Subdivision of the embryogenetic substrate into an array of cells, preparation for and occasionally even the initiation of the process of cell differentiation, or diversification and the creation of a cell aggregate (blastula) which through the activity of morphogenetic processes may be subjected to further morphological changes.
Patterns of cleavages
The pattern of cleavage due to organization of egg may be of the following types:
[I] Radial cleavage:
When the cleavage planes cut the zygote in such a manner that there appears a radial symmetry in the resulting blastomeres, the cleavage is called a radial cleavage.
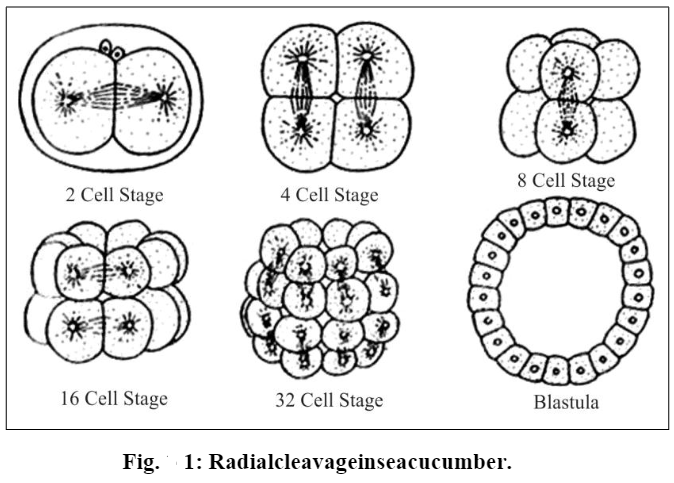
For example, for frog’s zygote divides by a vertical furrow into two equal blastomeres. The second cleavage furrow is also vertical but appears at right angles to the first. Thus, four blastomeres are produced. These four blastomeres remain sticking together. A horizontal cleavage then appears above the equatorial region to cut the four blastomeres into eight blastomeres with four smaller “upper” blastomeres and four bigger “lower” blastomeres. At the stage each bigger blastomere has a smaller blastomere sitting on its “head” and the blastomeres are arranged in four radial planes.
A blastula produced by radial cleavage can be cut along any meridian to get two identical halves. Radial cleavage is found in echinoderms.
[II] Biradial cleavage:
When the first three division planes do not stand at right angles to each other, the cleavage is termed as biradial. Examples are found in Polychoerus and Ctenophora.
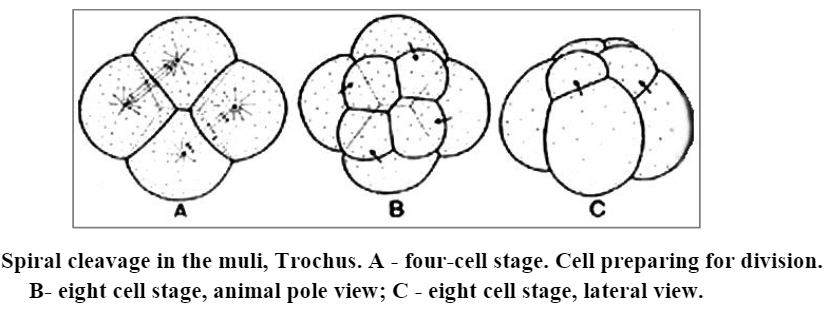
[III] Spiral cleavage:
The spiral cleavage is found in those forms in which there is a rotational movement of cell parts around the north pole-south pole axis of egg, leading to a displacement or inclination of the mitotic spindles with respect to the symmetrically disposed radii. Here, the cleavage planes are neither vertical nor horizontal but are slanting in relation to this axis. Moreover, each blastomeredivides to form one bigger cell (macromere) and a smaller cell (micromere). In such a cleavage the blastomeres of upper tier (micromeres) sits over the junction between each two of the vegetal blastomeres (macromeres). This is due to oblique position of the mitotic spindles. Therefore, it is also called oblique cleavage.
In successive cleavages the mitotic spindles are arranged in a sort of spiral. The turn of spiral may lie in a clockwise direction (dextral cleavage – right handed) or anticlockwise direction (left- handed-sinistral cleavage). Examples are found in Turbellaria, nematoda, rotifera, annelides and all themolluses except cephalopods.
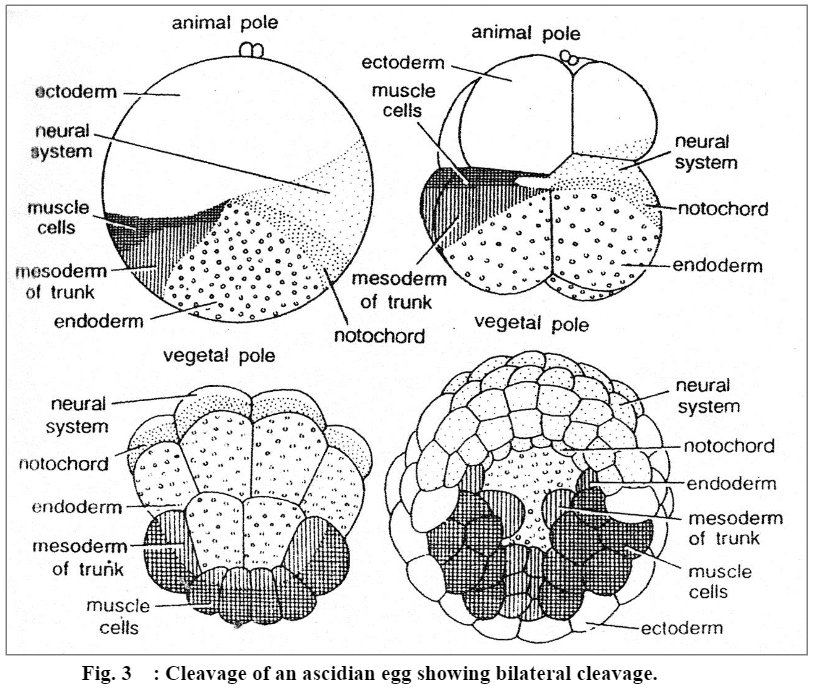
[IV] Bilateral cleavage:
In bilateral cleavage, the blastula can be cut vertically only along one plane to get two identical halves, the right and the left. Cleavage activity on one side is mirrored by activity on the other side. In most cases, the plane of bilateral symmetry is established by the plane of first cleavage furrow, which is bilaterally symmetrical. Examples are found in tunicates, Amphioxus, amphibians, and higher mammals.
[V] Asymmetrical :
According to the concept of potency. which refers to the total range of developmental possibilities, that an eggorablastomere is capable of realizing under any imposed conditions either natural or experimental, the following two types of cleavage have been recognized:
1) Determinate: The fertilized egg forms all the parts of the embryo by repeated division. Some eggs or ova have, even before cleavage, different regions earmarked to form different parts of the embryo. For example, in the ascidian eggs, the region will form the endoderm is fixed. If this region is dissected out from a fertilized egg, the embryo formed later will have no endoderm. Such eggs with predetermined regions are called mosaic (or determinate) eggs. Cleavages in mosaic eggs follow a precise pattern and each blastomere has its characteristic position and unalterable fate. Here cleavage separate different organ forming regions and are called determinate or mosaic cleavage. Examples are nematodes, annelids, molluscs and ascidians, which show determinate type of cleavage.
2) Indeterminate: In vertebrates the plan of cleavage is less rigid. Here the fertilized eggs do not have a predetermined region. If the region which normally forms the endoderm is removed from a fertilized sea urchin egg, the embryo formed later will still have the endoderm. Such eggs are called regulative or indeterminate eggs. In these eggs, as there are no predetermined regions and the cleavages cannot separate such regions, they simply cut the eggs into segments which have the potential of forming any organ. This type of cleavage is called indeterminate or regulative cleavage. Eggs of some groups of invertebrates and of all vertebrates show indeterminate cleavage.