Definition: It is a complex process. The union of the male and female gametes of same species is known as the fertilization. The fertilization is only method for the production of the diploid zygotes in vertebrates. In the process of fertilization, the haploid male gamete, sperm which carries the paternal genetic information, unites with the haploid female gamete, ovum, which carries genetic information of female parent, to form a diploid zygote. The zygote ultimately, produces a diploid multicellular organism by the several repeated and organized mitotic divisions and cellular differentiation.
Mechanism of Fertilization :
The process of fertilization completes in the following five stages:
(1) Encounter of spermatozoa and ova.
(2) Capacitation and contact.
(3) Acrosome reaction and penetration.
(4) Activation of ovum.
(5) Migration of pronuclei and amphimixis.
1) Encounter of spermatozoa and ova :
A major problem in sexual reproduction is how to bring together the spermatozoa and ova in the same locality in a fluid medium, so that individual sperms may reach the surface of ova at the right time. Previously it was thought that sperms are attracted towards the ripe eggs by chemotaxis. A chemical substance is found in the cortex of the eggs of sea urchin companularia, fishes etc. held responsible for the attraction of sperms to ripe eggs.
In general, most invertebrates, and vertebrates accomplish close approximation of spermatozoa and eggs through special devices or particular forms of behaviour. The primary needs for the encounter of spermatozoa and ova are fluid medium for the act of fertilization and delivery of large quantities of spermatozoa close to the numbers of ripe eggs at the right time.
According to the place and nature of fluid media following two kinds of fertilization have been reported:
1) External fertilization
2) Internal fertilization
1. External fertilization: This type of fertilization occurs in liquid medium outside the bodies of parent animals. Among fresh water animal. (fishes, amphibians and fresh water invertebrates), the timing of spawn of egg by the female and shedding of sperms by the male parent are very specific. As their spermatozoa remain active usually for a few minutes. sperms are delivered directly to the eggs of an individual female immediately after egg laying. But marine forms shed eggs and sperms freely into the surrounding water. The time interval between the laying of eggs and the shedding of sperms may even be weeks or months, because the saltish sea water serves as an important physiological medium for gametes.
During external fertilization, the movement of spermatozoa in a liquid medium is entirely at random and the spermatozoa collide with the as a matter of chance which occurs regularly in nature, partly due to enormous number of spermatozoa produced by the male and partly because the eggs being relatively larger targets, they can be hit by sperms fairly well.
2. Internal fertilization: In oviparous forms such as reptiles and birds, the eggs are completely enclosed in impermeable egg membranes or they are retained within the maternal body throughout development in ovoviviparous and viviparous animals. In all such cases the spermatozoa are delivered internally in the body of the female by some type of copulatory mechanism or by intromittent organ of the male. In such forms the fertilization may occur in the lower part of the oviduct (eg. Urodela) in the upper portion of oviduct (e.g. Salamenders, reptiles, birds and mammals) or in the ovarian follicles in viviparous fishes (e.g. Gambusiaaffinis) and eutherian mammals (e.g. Ericults).
In terrestrial animals, there is no problem of timing of the spawning of eggs and shedding of sperms because the mature sperms are commonly stored in a physiological medium capable of maintaining their life and potential activity for days or even for months, either in moisture conserving capsules or in compartments of male and female body to be picked up, transferred or utilized in one way or another. The movement of the spermatozoa from the site of deposition to the site of fertilization usually depends on the active swimming of the spermatozoa themselves transported passively by muscular contractions of the female tract and also by the counter currents in the cilia which propel backward flowing liquid content of the tract.
2) Capacitation and contract
In some animal species, the spermatozoa undergo a process called capacitation, before they are fully capable of fertilizing an egg of the same species or the change in spermatozoa which makes it capable of fertilizing the egg has been called capacitation. The phenomenon of capacitation relates to the specificity of fertilization, namely, the capacity of spermatozoa to fertilize eggs of the same species but not of other species.
Frank Lillie was the first to show that a chemical substance is discharged by the newly laid sea urchin egg. This substance diffuses into the sea water and causes sperm cells in the neighbourhood to become more active and attracted to the cell. This condition is called as chemotaxis, implying that the sperms were responding to this chemical substance. This substance has been found in the jelly which surrounds the eggs.
i) Agglutination. In most animals it has been observed that in the presence of ripe eggs or even the water in which ripe eggs of the same species have been lying for some time, the spermatozoa adhere to the surface of the egg by its lateral side and even to each other. This reaction is usually visible within a few seconds and the spermatozoa are seen to clump together head-to-head or less commonly tail to tail. This adhesion of spermatozoa results in their clumping or agglutination. It depends to a large extent on the environmental conditions.
ii) Fertilizin-antifertilizin reaction. The cause of agglutination of spermatozoa was studied by Lillie (1919). He observed that fertilizin and antifertilizin occur in the eggs and sperms respectively. They are directly involved in the reaction between the egg and the spermatozoon. The main source of fertilizin is the egg itself and it is located in the plasma membrane. However, in the eggs of sea urchin and other echinoderms, it is produced by the layer of jelly surrounding the egg, and becomes accumulated in the external gelatinous coat.
The fertilizin is a gel formed of glycoprotein or mucopolysaccharide. As a protein, it contains a number of amino acids and as a polysaccharide it includes molecules of one or more monosaccharides. The monosaccharides glucose, fructose, fucose or galactose are esterified by sulphuric acid as shown in the formula in fig.1.
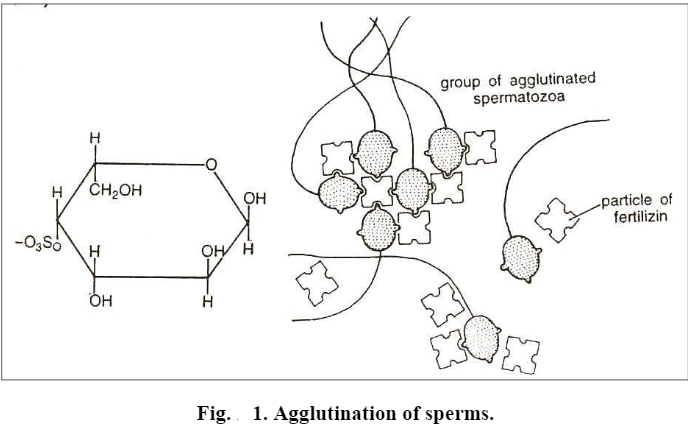
Both the amino acids and polysaccharides vary from one species to another, that is why each species possesses its specific type of fertilizin.
The molecules of fertilizins are quite largethe molecular weight is about 300,000 and each molecule may have more than one “active group” so that one fertilizin particle may become attached to two or more spermatozoa thus binding them together.
The surface layer of cytoplasm of spermatozoa (i.e. sperm plasma membrane) contains another species specific acid protein known as antifertilizin(or antifertilizins). The antifertilizins can be extracted from the spermatozoa by heating, freezing and thawing, or acidifying the water.
The remarkable peculiarity of the fertilizins and antifertilizins is that they combine in a specific manner; that is, the egg fertilizin of a particular species reacts best with the sperm antifertilizin of the same species. Reactions between the species which are nearly related to each other are very much weaker. The reaction between fertilizin and antifertilizini much similar to the reaction between antigen and antibody. In both cases a “chemical lock” is formed between two complementary substances Thus during capacitation and contact stage of fertilization when spermatozoa and eggs of same species come in physical contact of each other, a chemical lock is established between the antifertilizin molecules of spermatozoa and fertilizin molecules of unfertilized egg and due to the fact, many spermatozoa adhere to the surface of an unfertilized egg.
It has been suggested that the main function of fertilizin-antifertilizin is to thin out the number of spermatozoa around the egg, so that the chances of two or more spermatozoa fusing with the egg at the same time are diminished.
3) Acrosome reaction and penetration
As mentioned earlier, except porifers and coelenterates, where the surface of the ripe egg is seldom naked most animal eggs are enveloped by one or more egg membranes or gelatinous layers or follicle cells or both, outside the plasma membrane. These layers constitute barriers for the penetration by spermatozoa and serve in preventing fertilization by more than one spermatozoon or by sperm of other species.
When a spermatozoon is attached to surface of the egg, it becomes motionless. Its penetration through egg membranes and also through the plasma membrane of the egg is achieved by some physiochemical activity of the sperm acrosome. Certain enzymatic proteins called sperm lysins are produced presumably by the sperm acrosome.
The sperm lysins differ from one animal group to another. In some cases, the dissolution of the egg envelopes may be brought about by simpler means. Thus, it is believed that the jelly coat of echinoderm eggs may be dissolved as a result of acidification of seawater by carbon dioxide produced by the spermatozoa in the course of their respiration. In the case of eggs with very thick and resistant envelopes, such as the egg envelopes of fishes and insects, the sperm cannot reach the egg at all points but must penetrate through a special canal, the micropyle, left in the egg envelope, the chorion. In mammals, when the eggs are released from the ovary, they are commonly encased in a layer of follicular cells, called corona radiata. These cells are held together by an adhesive cementing substance called hyaluronic acid, a mucopolysaccharide. The corona radiata, thus, acts as a barrier through which the spermatozoon must first penetrate to reach the plasma membrane of the egg. For this purpose, the sperm’s acrosome produces an enzyme, hyaluronidase, which serves to dissolve the adhesive substance and disperses the cells of corona radiata. The breaking of the membranous barriers is not only mediated by lytic agents provided by the acrosome of the spermatozoon, but the acrosome itself undergoes morphological changes and forms acrosomal filament which helps the sperm penetration into the egg interior. The entire process has been well illustrated in echinoderms, annelids and Saccoglossus.
Acrosome reaction and penetration in Saccogiossus. Penetration and acrosomal reaction of spermatozoa of Saccoglossus has been best described by Colwin and Colwin (1967). A spermatozoon of Saccoglossus has spherical nucleus, a flat tail and an acrosomal vesicle at the forwarding end of the sperm head.
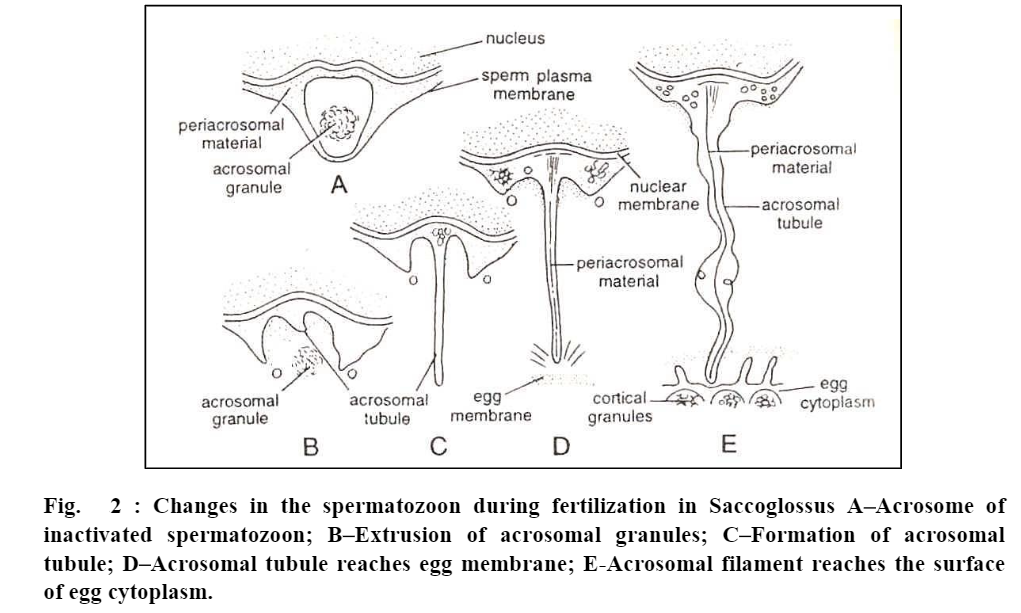
The acrosomal vesicle is bounded by an acrosomal membrane and contains a large, dense acrosomal granule. The granule is surrounded in large part by fine, grainy material except at the apex where an apical space lies between the granule and membrane. The space between the acrosomal membrane and sperm plasma membrane and also, the space between acrosomal membrane and nuclear membrane, are filled by some material called periacrosomal material.
As the spermatozoon of Saccog/ossus makes its initial contact with the egg envelope, the following events occur.
(a) Bursting of acrosome. Apex of the acrosome bursts so that the membrane of sperm and acrosome open apically and consequently expose the interior of the acrosomal vesicle to the outside. The two membranes join around the margin of the opening.
(b) Release of lytic enzymes. Acrosomal granule is released and comes in contact with the egg envelope. It contains lytic enzymes which make passage through the egg envelopes. Therefore, shortly after its release, the acrosomal granule disintegrates and disappears.
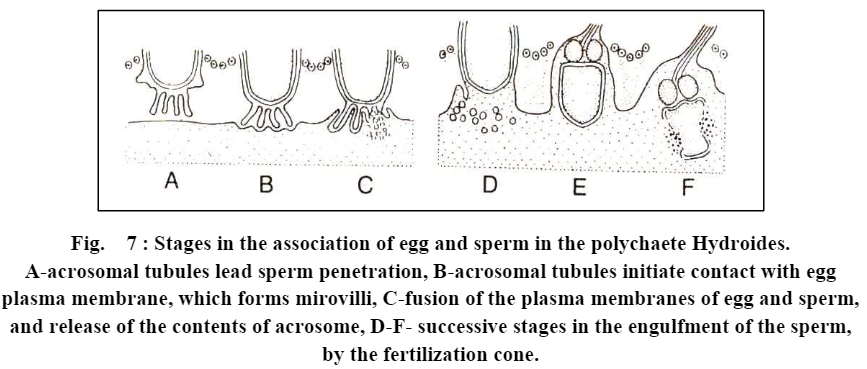
(c) Formation of acrosomal tubule. Shallow depression of the acrosomal membrane close to the nucleus now deepens and soon lengthens into a long slender acrosomal tubule. The tubule becomes twice as long as the sperm nucleus.
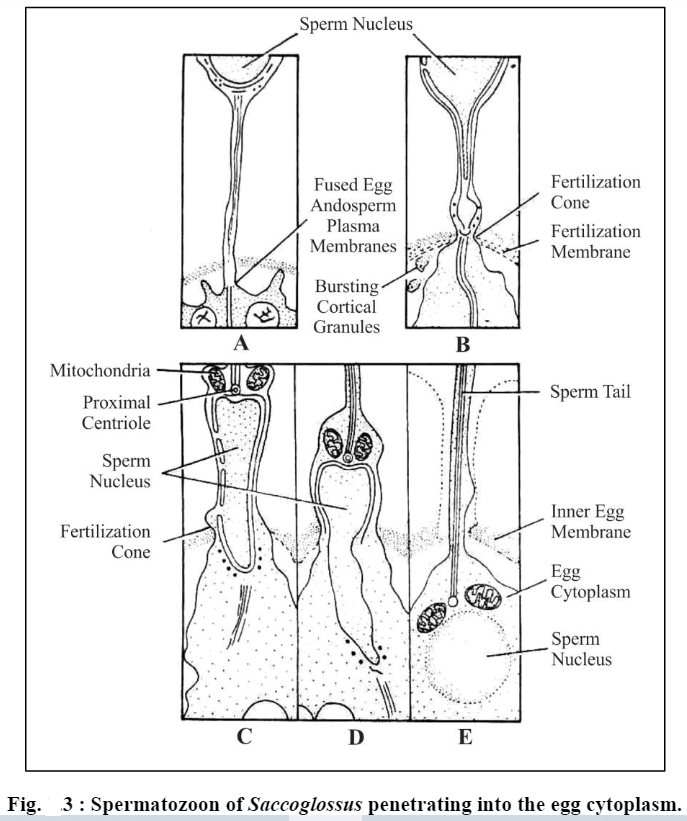
(d) Eversion of the acrosomal membrane. Rest of the acrosomal membrane everts and is added to the acrosomal tubule at its base. This is simply an unfolding of the already continuous membrane.
(e) Fusion of acrosomal tubule with egg membrane. Acrosomal tubule gradually enters through the passage of egg envelope, which has been made previously by acrosomal lytic enzymes, and ultimately touches and fuses with the egg plasma membrane.
(f) Passage of sperm contents. Fertilization cone protrudes from the egg and engulfs acrosomal tubule. The nucleus of spermatozoon is drawn out towards fertilization cone. The acrosomal tubule dissolves: elongated nucleus along with middle piece of spermatozoon is engulfed into the egg cytoplasm.
In other animals, similar types of events have been observed during the penetration of sperm into the egg envelope, i.e. before penetrating the egg contents and before activating the egg for further developmental events the spermatozoa of many animals themselves get activated like the spermatozoa of Saccoglossus. In them sperm activation includes rupturing of acrosome and formation of acrosomal filament or acrosomal tubule. However, the number and size of acrosomal tubules may vary from species to species, as some annelids (e.g., Aidroideshexagonus) have several acrosomal tubules.
The mammalian spermatozoa though possess acrosome do not develop acrosomal filaments. The spermatozoon appears to contact the surface of the egg by its lateral aspect. Following this action, the plasma membrane of the egg and the spermatozoon dissolve at the point of contact, and the spermatozoon is drawn into the interior of the egg.
4) Activation of ovum
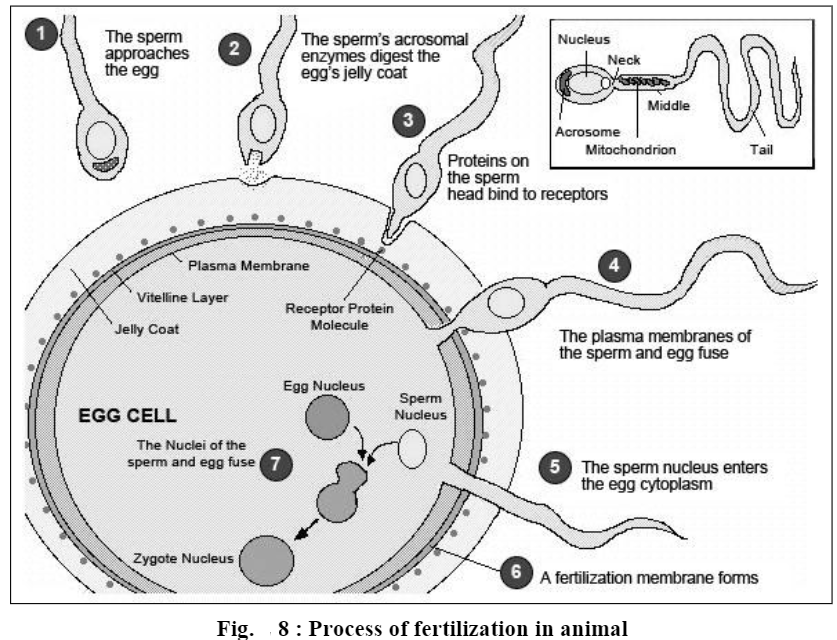
Activation of ovum is that aspect of fertilization by which an egg is released from its inactive state and begins to develop. As soon as the apex of acrosomal tubule of a spermatozoon touches the surface of egg plasma membrane fusion of both membranes (i.e. plasma membranes of sperm and egg) over this limited area of contact takes place and a single continuous mosaic membrane is formed. Thus, the plasma membrane of both gametes (sperm and ovum) becomes continuous and forms a single cell, called zygote. At this very time, certain very important changes occur in the cytoplasm of egg:
a) Fertilization cone formation.
b) Cortical reactions and formation of fertilization membrane.
c) Metabolic activation.
a) Fertilization cone formation. Immediately after the acrosomal filament of spermatozoon touches the surface of the egg, the cytoplasm of the egg bulges forward at the point of contact, producing a process of hyaline cytoplasm called the fertilization cone.
Fertilization cone develops or appears in many forms. It may be in the form of a more or less simple conical protrusion or it may consist of several irregular pseudopodium-like processes, or in some cases it may take the form of a cytoplasmic cylinder stretching forward along the acrosomal filament or tubule. Whatever its shape, the fertilization cone gradually engulfs the spermatozoon and then begins to retract. Here one point should be clear, that normally the spermatozoon does not enter the egg cytoplasm intact, nor, it is swallowed but the sperm nucleus and other sperm structures (periacrosomal material, proximal centriole and mitochondria of mid-piece of spermatozoon) pass to the fertilization cone of the egg. The plasma membrane of sperm becomes one entity of plasma membrane of the egg.
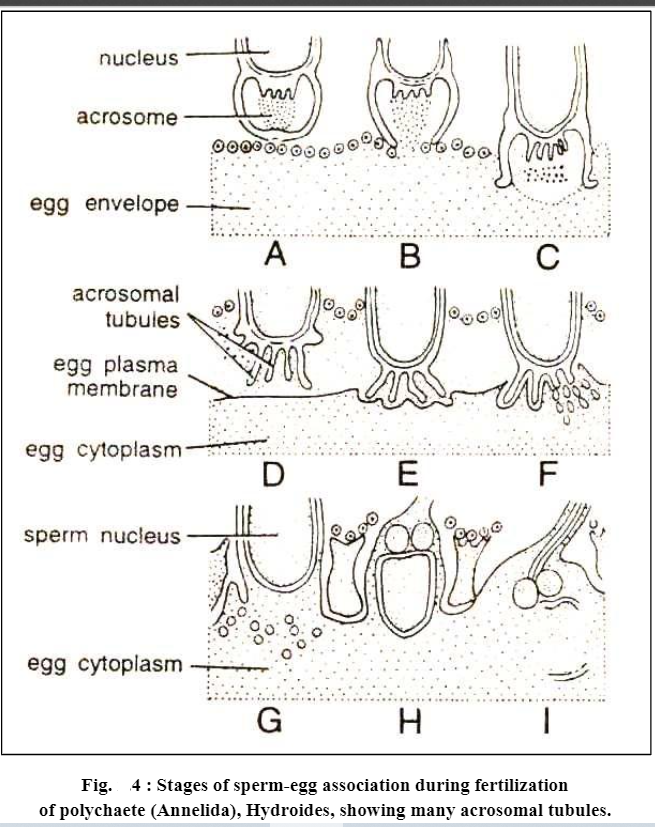
Further during the whole penetration process of sperms into the egg, acrosomal granule never makes its entry into the egg, but only the periacrosomal material is injected into the egg cytoplasm along with other contents of the sperm. Some workers suggest that this periacrosomal material is responsible for the activation of egg.
There exists some variation in different animals, as to how much of the spermatozoon is taken into the interior of egg during fertilization. In mammals, complete structures of spermatozoon (viz., nucleus, mid piece, tail etc.) penetrate into the egg cytoplasm. In echinoderms, sperm tailremains exterior to the vitelline membrane; in Neries only sperm head and proximal centriole enter the egg cytoplasm. In most animals, however, the sperm nucleus and mid piece make their entry into the egg as a rule.
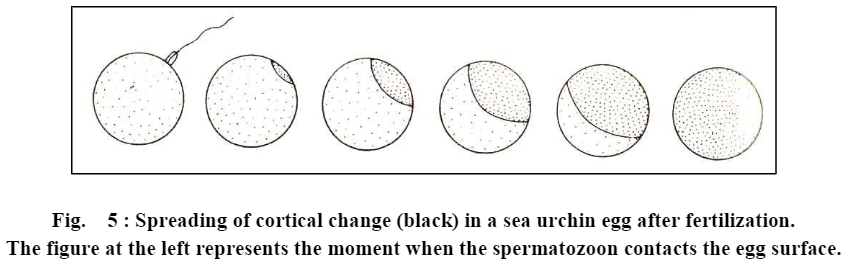
b. Cortical reactions and formation of fertilization membrane. Even before the fertilization cone is formed and the spermatozoon penetrates into the interior of egg, a chain of physico-chemical reactions is set in the egg cortex. All these reactions are collectively called cortical reactions. These reactions may differ from one group of animals to another, but in most groups, the cortical reactions lead to the formation of fertilization membrane outside the egg plasma membrane. This membrane blocks the entry of the late arriving spermatozoa in the egg interior, and thus avoids polyspermy. The process of cortical reactions and fertilization membrane formation in different groups of animals is as under:
(a) Sea urchins. In sea urchins, as soon as the apical end of acrosomal tubule touches the surface of egg, from the site of contact, a wavelike colour change from yellow to white (under dark field microscopy) travels rapidly around the egg cortex and is shortly followed by the elevation of fertilization cone from the egg surface and the formation of fertilization membrane around the egg plasma membrane.
Electron micrographs of sea urchin’s unfertilized eggs show that the egg cortex is bounded by two membranes (i) an outer 30 A° thick, vitelline membrane, and (ii) an inner, 60 A° thick plasma membrane.
Beneath the plasma membrane occurs a layer of cortical granules. A fertilization membrane is formed in the following stages:
The outer vitelline membrane separated from the plasma membrane undergoes expansion and becomes the outer layer of the fertilization membrane. The cortical granules explode and release the following three components:
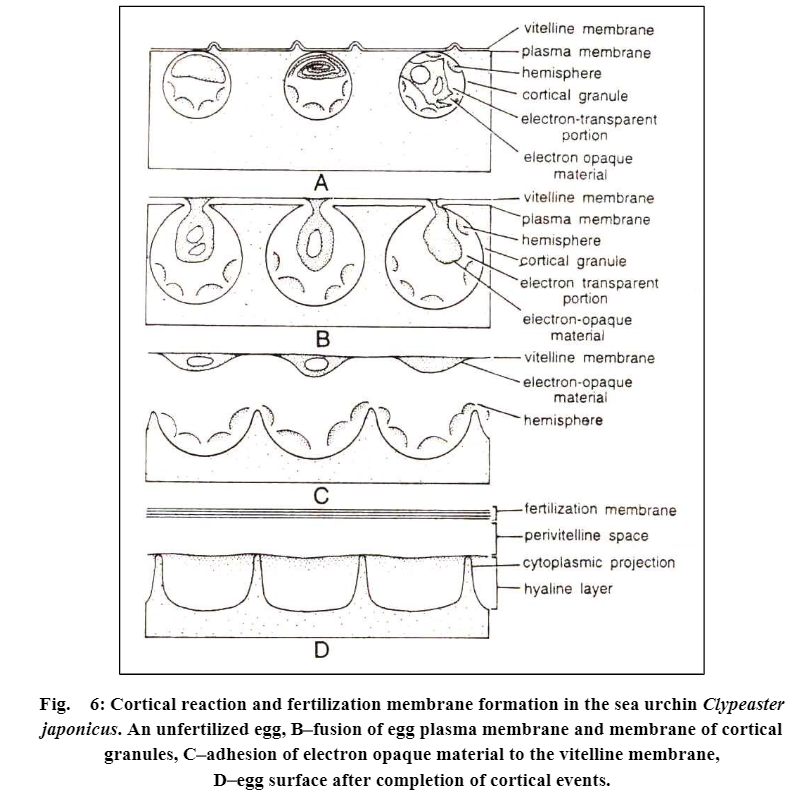
(1) Dark, denser, lamellar and folded parts of the granules-these lamellar bodies unfold and fuse with the inner side of the already elevating membrane, the vitelline membrane.
(2) Globules, which fuse together and build up a new surface of the viscous hyaline layer. just at the outer side of the egg plasma membrane. The hyaline layer adheres closely to the surface of the egg and during cleavage, it helps to keep the blastomeres together.
(3) The liquified component of the cortical granules fills the perivitelline space between the new egg surface and now the completed and elevated fertilization membrane. It contains mucopolysaccharides and abundant water.
All these structures, namely, vitelline membrane and contents of cortical granules, thus form a fertilization membrane, which is much thicker (upto 900 A°) and stronger.
Among other invertebrates, fertilization membrane formation has been observed only in certain annelids (e.g., Nereis). In others no structural but metabolic changes have been observed in egg cortex.
(b) Vertebrates. In vertebrates the changes which occur in the cortex are similar to sea urchins. In bony fishes and frog, the cortical granules are broken down immediately after sperm’s penetration into the egg cytoplasm. Their contents become liquified and extruded on the surface of the plasma membrane of the egg and fill the perivitelline space occurring in between the chorion and plasma membrane and in between the vitelline membrane and egg plasma membrane in the frog. In both cases, the vitelline membrane or chorion itself does not transform into the fertilization membrane as in sea urchins. The chorion in fishes becomes hardened or “tanned” after fertilization and no new membrane is formed in either of the two animals.
In some mammals (man, rabbit, etc.) the cortical granules burst and release their contents into the perivitelline space created between the egg plasmalemma and the zonapellucida.
In urodele amphibians and some mammals, which lack cortical granules, neither any cortical reaction nor fertilization membrane formation occurs.
c) Metabolic activation. After the sperm penetrates the unfertilized eggs a series of diverse cytoplasmic reactions are initiated. Following metabolic changes occur in the egg during fertilization.
(i) Changes in plasma membrane. The permeability of plasma membrane increases for the molecules of water and certain other chemicals like ethylene glycol, phosphate, K+ ete. At fertilization, the electrical potential of plasma membrane becomes more positive and it gradually becomes more negative. The change in the potentiality of the membrane is governed by the unequal distribution of chloride ions.
Besides this, a plasma membrane enzyme adenylcyclase becomes activated at the time of fertilization and it starts the formation of a chemical molecule 3′ – 5′ cyclic AMP, which is supposed to activate most of the metabolic reactions in a fertilized egg.
(ii) Tonic changes. Certain intracellular changes occur in the concentration of cations, especially those of sodium, potassrum and calcium. The change in calcium ion concentration in a fertilized egg has great significance in the metabolic activation of the egg.
(iii) Changes in the rate of respiration. In a fertilized egg, the rate of respiration either increases (e.g. sea urchins) or decreases (e.g. Chaetopterus and Molluscs, Cumingia) or remains static (e.g. Bufo and Fundulus). There appears to be a relation between the post- fertilization oxygen consumption and the stage of maturation of the egg at fertilization. Because at the time of fertilization the sea urchin egg has completed maturation, the egg of Bufo is at second maturation division stage and the egg of Chaertopterus is at the first maturation division stage.
The increased oxygen consumption is related with the oxidation of glycogen and other food stuffs of the egg and synthesis of numerous ATP molecules.
(iv) Co-enzyme changes. The primary action of the spermatozoon consists of the release or activation of the oxidative enzymes of the egg and that the ensuing increase in oxidation provides the energy necessary for the performance of other changes in the egg and for the development of the egg in general.
In a fertilized egg inter conversion of pyridine coenzyme, NAD into another co- enzyme NADP and also NADPH due to phosphorylation of the NAD in the presence of a enzyme NAD Kinase takes place.
![]()
There are ample evidences that NAD kinase enzyme although present in the unfertilized egg, is in inactive state. It is activated only at the time of fertilization. The increased NADP and NADPH contents may initiate many synthetic pathways of fertilized egg.
(v) Changes in the rate of protein synthesis. The cytoplasm of a mature unfertilized egg contains complete machinery for protein synthesis, such as DNA molecules, tRNA, mRNA, ribosomes and proteolytic enzymes required during protein synthesis, but none or very little protein synthesis occurs because the mRNA of unfertilized egg remains “masked”. There are evidences that during later phases of oogenesis some inhibitor or repressor proteins are manufactured in sea urchin’s egg which inactivate chromosomal genes, mRNA molecules, ribosomes etc. During fertilization there is an increase in proteolytic activity of the egg immediately following the penetration of spermatozoon which removes these inhibitor proteins from them and unmasks the mRNA and active protein synthesis is started. In the egg of frog, however, the rate of protein synthesis is increased quite early at the stage of ovulation itself.
(vi) Initiation of mitosis. The initiation of mitosis for cleavage is the most significant aspect of egg activation. The initiation of mitosis occurs because (i) the rate of DNA synthesis increases with great pace immediately after fertilization; (ii) the unfertilized egg cytoplasm although possesses a centriole, yet this centriole is incapable of division and also to form a mitotic spindle. Thus, sperm stimulates the first mitotic division (cleavage) of fertilized egg by contributing its centriole to the egg. In some animals after fertilization, considerable amount of acid is formed during first several months.
Significance of fertilization:
1. The entrance of sperm activates the secondary oocyte to complete its second maturation division.
2. Induces the movement of cytoplasm and the pigment granules and marks the fertilization track.
3. Induces the second centriole and initiates spindle formation.
4. Restores the diploid number of chromosome and recombines the maternal and paternal genetic traits.
5. Induces change in the periphery of egg and this change precludes the entry of other sperms.
6. This causes separation of vitelline membrane which allows the rotation of egg inside.
7. This enhances the metabolic activities and thus renews the vigour and vitality.
8. The amount of cytoplasm lost due to the separation of polocytes is restores.
9. Establishes a new plane of egg axis.
10. Causes rearrangement and organization of egg cytoplasm into distinct organ forming areas.