Muscle cells
- Muscle cells are specialized for contraction: every muscular function, from running and jumping, to ventilating the gills or lungs, propelling blood through the vascular system or food through the gut, or expelling an egg from the oviduct, is carried out by muscle cells working in concert.
- Many protoctistans have contractile stalks (e.g. the ciliophoran Stentor sp.), and contractile cells occur in sponges associated with the openings (ostia) through which water is discharged into the spongocoel central cavity.
- Contractile muscle cells are found in the column walls of Cnidaria such as sea-anenomes. In higher animals, single muscle cells persist such as the myoepithelial cells associated with mammalian sweat glands. Most contractile cells in Metazoa are organized and differentiated into muscular tissue.
Properties of Muscle
Muscles generate force. In humans they comprise 40–50% of the total body weight. Muscle is:
- excitable: it receives and responds to stimuli;
- contractile: it shortens and thickens to do work;
- extensible: it can be stretched passively when relaxed;
- elastic: it returns to its original shape after contraction and extension.
Note that muscles only do work when they contract: contraction is the active process.
Most muscles associated with the skeleton are arranged in opposing, antagonistic pairs: when one contracts the other is passively stretched (e.g. the biceps and the triceps bending and straightening the human elbow). In bivalve molluscs, the closing adductor muscles work against the elastic spring ligament in the hinge of the shell.
Functions of Muscle
There are three major functions for muscle:
(1) motion, including locomotion;
(2) maintenance of posture;
(3) heat generation (particularly in homoiotherms): approximately 85% of human body heat is generated by muscle contractions; brief involuntary muscle contractions (shivering) generate heat when the animal is cold.
Muscle types in Mammals
There are three types of muscle tissue.
(1) Striped (striated) muscle moves the skeleton; often called voluntary (because its movement is usually voluntary) or skeletal muscle.
(2) Cardiac muscle is a special type of striped muscle in the walls of the heart; contraction is involuntary.
(3) Smooth muscle surrounds the walls of internal organs such as the gut, bladder, blood vessels and the uterus; contraction is involuntary.
Striped muscle
- The disposition of striped (skeletal or striated) muscles and their names is best studied using an illustrated guide. Striped muscles are surrounded by superficial fasciae composed of connective and adipose tissues: they provide a route for nerves and blood vessels.
- Deep fascia is connective tissue which holds muscles together and separates them into functioning units. The entire muscle is also wrapped in fibrous connective tissue: the epimysium.
- Bundles of muscle cells (fasciculi) are covered by perimysium. Endomysium penetrates into the fascicles and surrounds and separates the muscle cells.
- The epimysium, perimysium and endomysium contribute collagenous fibers to tendons which attach muscle to the periosteum of bone.
- Tendons can be flat or broadly cylindrical. Muscles are richly vascularized to deliver oxygen and glucose and to remove waste products; they are also extensively innervated.
- The muscle fibers (or cells, bounded by a plasma membrane or sarcolemma) contain contractile organelles called myofibrils ; each has a bundle of myofilaments divided lengthways into repeating structural units called sarcomeres.
- The sarcomeres are divided into bands of different densities, giving the striped appearance characteristic of this muscle type.
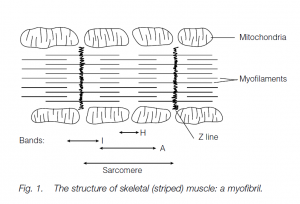
There are two kinds of myofilaments:
- heavy myosin filaments;
- light actin filaments.
X-ray crystallography shows that, in cross-section, the filaments line up in hexagonal array. Actin filaments are held by their attachment to the Z lines (made up of the protein α-actinin). I bands are regions containing actin filaments alone; the light H band in the middle of each sarcomere is where only the heavy myosin filaments are present; the myosin and actin filaments overlap in the dark A bands.
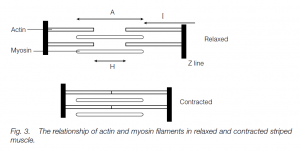
Sliding filament Model
- When a striped muscle contracts, the filaments (which do not change length) slide past each other. In each sarcomere, the many globular myosin heads which project laterally along each end of the heavy myosin filament attach to the actin filament and change conformation.
- The myosin pulls at the actin filaments adjacent to it. The myosin heads have been energetically charged, adopting a conformation in which they can bind to actin.
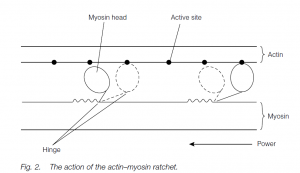
- This binding elicits the conformational change that provides the force for filament sliding and exposes an ATP-binding site.
- ATP binding causes an allosteric (shape) change that promotes detachment of the head from actin. Dephosphorylation of ATP provides the energy to re-establish the actin binding; thus the process is repeated many times (each using one ATP molecule) and the myosin pulls along the actin filament in a ratchet fashion.
- Since the ends of each myosin filament pull in opposite directions, towards the sarcomere center, the myosin pulls the two actin regions closer and with them the Z lines: thus the whole muscle contracts. (Actin–myosin systems are found elsewhere: e.g. pulling chromosomes apart at mitosis.)
Energy for Contraction
- The energy for contraction is provided by ATP: the myosin heads have an ATPase, the active site of which is exposed upon actin binding.
- The ATP synthesized and stored in muscle can only sustain a few seconds of vigorous exercise. Striped muscles contain high levels of phosphocreatine: this breaks down to release energy [some of which is used to make more ATP from adenosine diphosphate (ADP) and the phosphate is released]:
Longer-term energy needs must be supplied by cellular respiration:
- breakdown of glycogen (via glucose) to pyruvate with the synthesis of ATP, used as above: this is glycolysis and does not require oxygen;
- oxidation of pyruvate, in the presence of oxygen, to yield carbon dioxide, water and large amounts of ATP (38 molecules of ATP following the total oxidation of one glucose molecule: this is aerobic respiration).
- In the absence of oxygen, the pyruvate is normally converted to lactate: this is anaerobic metabolism. In heavy exercise, the lactate spills over into the blood.
- In humans, the glycogen/lactic acid system can provide energy for about 40 seconds of maximum muscle activity. The lactate constitutes an ‘oxygen debt’ which must later be paid off by oxidizing the lactate to carbon dioxide and water.
- Aerobic respiration can continue more or less indefinitely, provided oxygen and glucose (or another fuel such as fats) are available.
Regulation of Contraction
- Regulation of contraction is effected primarily by the proteins tropomyosin and troponin. Tropomyosin blocks the actin sites to which myosin will attach.
- Troponin has sites which bind calcium: this changes the troponin shape, so allowing the troponin to displace (‘push aside’) the tropomyosin in order that actin and myosin can interact.
- Neuromuscular transmitter release (acetylcholine from the endings of motor neurons innervating striped muscle in mammals) triggers an action potential in the plasma membrane (sarcolemma) of the muscle fiber (cell).
- The T (transverse) tubule system is continuous with the cell membrane (sarcolemma), and runs through the muscle fibers close to the Z lines. Thus action potentials can be rapidly transmitted into the fibers.
- The sarcoplasmic reticulum is a second system which encloses the myofibrils. It contains calcium ions, and by sequestering them it keeps the concentration around the myofibrils low.
- The reticulum expands near the Z lines of each sarcomere where the reticulum is in intimate contact with the T tubule system (although the two systems are not physically continuous).
- The action potential is transmitted along the membranes of the T tubules (propagation is similar to that along the plasma membrane of a neuron) and stimulates calcium release from the sarcoplasmic reticulum, so that the intracellular level of calcium increases 100-fold; the calcium initiates contraction by binding to troponin.
- Immediately after the action potential has activated the system, the calcium is pumped back into the sarcoplasmic reticulum (an ATP-dependent process); acetylcholinase (or acetylcholinesterase) breaks down the neuromuscular transmitter acetylcholine.
- Each contraction lasts for a few tenths of a second, until the calcium pumps have reduced the intracellular calcium pool to a point where the contractile apparatus is no longer operational; repeated action potentials cause long-term contraction or muscle tonus or tetanus.
Cardiac muscle
Cardiac muscle resembles striped muscle in that it has similar assemblies of actin and myosin and has a striated appearance. The fibers are shorter and have branched ends. Contraction is spontaneous and involuntary. Gap junctions in the intercalated disks that join the branched cells longitudinally transmit action potentials so that the whole of the muscle contracts synchronously.
Smooth muscle
Smooth, involuntary muscle is also made up of actin and myosin, but the molecules are arranged much less regularly so there are no striations (thus ‘smooth’). Actin–myosin interactions are similar to those in striped muscle, although the actin filaments are attached to dense bodies rather than to Z lines. Calcium is not stored in a sarcoplasmic reticulum, but in the extracellular fluid. Contractions tend to be slower and more prolonged. In hollow organs such as the gut, the muscle fibers are arranged in sheets with a longitudinal orientation outside and a circular orientation inside.
In vertebrates, smooth muscle fibers are divisible into two major types.
(1) Multi-unit smooth muscle fibers (e.g. in the iris of the eye): here nerve impulses reach each fiber from a motor nerve end-plate.
(2) Visceral smooth muscle fibers (e.g. gut and uterus walls): here only a few fibers have motor nerve end-plates, the action potential flowing to adjacent fibers through gap junctions. Such muscle fibers will contract spontaneously when stretched.
Muscle contraction physiology
- For most vertebrate striped muscles, a motor neuron branches so that there are several motor end-plates each on one of several muscle fibers (cells): the neuron plus its fibers is the motor unit.
- A nerve impulse causes all fibers in the unit to contract simultaneously. The more fibers per unit, the less the control which can be exercised (some eyeball muscles may have only two or three fibers per neuron while locomotory muscles may have 400–500 fibers per neuron).
- Contraction of a fiber may be initiated by a single nerve impulse, or several impulses with a summating effect may be necessary.
- There is a latent period of a few milliseconds between the arrival of the impulse and actin–myosin interaction. Tension develops during the contraction period and then decreases during the longer relaxation period.
- If a muscle works against a constant load so that the muscle shortens, this is isotonic contraction (e.g. lifting this book from the desk) or positive work; if the muscle pulls against an immovable object (e.g. pulling at a locked door), this is isometric contraction: internal shortening and tension develop as the elastic elements in the muscle are stretched but the muscle does not shorten.
Phasic and tonic Muscles
Most striped muscles in vertebrates are phasic (twitch) muscles. Each fiber has one motor nerve end-plate. When the motor unit is stimulated by its neuron, a single stimulus with a threshold intensity leads to a twitch which is all-ornothing because the action potential spreads rapidly throughout the whole fiber. Gradation in muscle tension is facilitated by:
- imposing a second twitch on the first before the latter’s effects have decayed (not all the myosin heads are activated during the time available for the first twitch) this is temporal summation; or
- delivering impulses to reach the motor unit so rapidly that no relaxation can occur between action potentials, so eliciting a plateau of tension or state of tetanus which persists until the impulses cease or the muscle fatigues; or
- progressively recruiting more motor units to contract: this is spatial summation. Some muscle fibers (but not the same ones continuously) will always be contracted in a given muscle: this is called tonus and allows the muscle to be held firm.
Slow phasic fibers are used to maintain posture and in endurance activities: they are often rich in myoglobin and are dark red in color. Such fibers are rich in mitochondria. Their slow contraction allows complete, aerobic oxidation of fuels and they fatigue only slowly. Fast phasic (glycolytic) fibers are used for quick bursts of movement; they lack myoglobin and have fewer mitochondria; their metabolism is often anaerobic. The two fiber types may be mixed (as in most mammalian muscles) or separated (the red and white meat in chicken or herring).
Tonic muscles are like phasic muscles with slow fibers, except that they have many nerve end-plates per fiber and do not follow the all-or-nothing phenomenon seen in a phasic muscle fiber twitch. Action potentials do not spread far and gradation in contraction is effected by more frequent impulses. Bivalve (e.g. oyster) adductor muscles contain phasic and tonic fibers, the former snapping the valves of the shell closed while the tonic fibers keep it closed.
Arthropod Muscles
Arthropod muscles (e.g. in crab claws) have very few neurons per muscle, and a given neuron may innervate several muscles. Each fiber may have multiple end-plates from each motor neuron, and up to five neurons may innervate one fiber: at least one neuron is inhibitory whereas the others facilitate varying degrees of slow or fast contraction. Combinations of motor stimulation can thus permit a gradation of action.
Muscle work and Power
Muscle force reflects the number of actin–myosin cross-bridges formed, in turn dependent on the number of fibers within the muscle, roughly proportional to its cross-sectional area: Force a Cross-sectional area of the muscle Force of contraction per unit cross-sectional area is usually between 4 and 6 kg cm–2. Muscles produce most force when contracting around their resting length.
Work is the product of force and the distance through which the force works (force × distance): it is more or less constant per gram of muscle, but large muscles will, of course, generate more force. In some muscles, the fibers are arranged longitudinally (fusiform pattern) which permits contraction over a greater distance compared with muscles with the pennate or feathered pattern of fibers where the fibers are arranged to allow shorter fiber contraction, but the larger number of fibers generate much more force. Thus large muscles (e.g. the quadriceps of the human thigh), with a large cross-sectional area and therefore force, which can contract significant distances (e.g. about one-third the resting length), can do large amounts of work compared with small, short muscles.
Power is the rate of doing work. Very small muscles (e.g. those of the eyeball in a small rodent) are very powerful in that they contract very quickly: the power output per unit weight of muscle is higher than that of the equivalent, larger muscles of a larger mammal. Muscles develop their maximum power at intermediate velocities: the actual velocity depends on whether the fibers are phasic or tonic, but efficiency is at a maximum at 20–30% of maximum velocity. Energy is lost because of inefficiencies in the energy interconversion processes: much will be lost as heat and to overcome internal friction in the muscles
Evolutionary Aspects
Somatic muscles in vertebrates are associated with the body wall and appendages (fins and limbs) and are usually striped; visceral muscles are associated with the gut and are usually smooth. Fish somatic muscles mainly comprise segmental myomeres which facilitate the undulation of swimming. Paired fins are moved by a dorsal abductor muscle (pulling the fin up) and a ventral adductor muscle (pulling it down). On land, the segmental arrangement of muscles is largely lost. The fin abductor and adductor muscles become divided to form the limb muscles attached to the limb girdles, and additional muscles develop in the limbs themselves.