Nitrogenous Excretion
Nitrogenous excretion is the removal of nitrogenous waste substances of metabolism.
Nitrogenous waste substances
During protein catabolism (breakdown) amino acids are deaminated to form a keto-acid and ammonia. Ammonia is very soluble in water and, as it goes into solution, some combines with protons in the water to form NH4+. This is very toxic, but is quickly flushed from the body if copious water is available (500 ml of water will flush away 1 g of nitrogen). Most fishes excrete ammonia as do amphibian tadpoles. Terrestrial animals need to conserve water so they detoxify ammonia to urea in the liver. This saves about 90% of the water needed to void 1 g of nitrogen. Urea is the main nitrogenous excretory product of mammals.
Further detoxication of ammonia yields the purine uric acid, the principle nitrogenous waste product of land snails, insects, reptiles and birds. This only requires less than 10 ml water to eliminate 1 g of nitrogen, but uric acid is metabolically expensive to produce.
Invertebrate excretory organs
Protoctistans use a contractile vacuole. In Paramecium sp. there is one at each end of the ciliate, each composed of a radiating ring of tubules which deliver water to a central vesicle; on reaching a certain size, the vesicle discharges (by fibrils contracting and then by hydrostatic pressure) through a pore in the outer pellicle.
In Metazoa a few animals void ammonia by simple diffusion to the exterior (e.g. cnidarians). Most animals employ excretory organs which can be tubular or saccular. In tubular organs, the excretory tubule opens into the coelom. Fluid is filtered from the blood into the coelomic fluid whence it passes into the tubule: here selective resorption of salts, sugars, amino acids and water may occur, and secretion of wastes by the tubule wall takes place, helped by the blood vessels adjacent to the tubule. The final solution produced is urine.
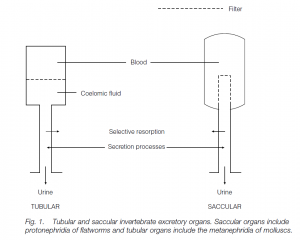
Saccular organs have a closed tubule which does not open into the coelom: the tubule receives filtrate directly from the blood. The filtrate composition is modified by absorption and secretion to yield urine. In many animals the osmoregulatory /excretory tubules are termed nephridia, which open to the exterior by a nephridiopore. Flatworms and some marine annelids have protonephridia. The inner, blind end of this saccular organ has a terminal cell with ‘flickering’ cilia, a flame cell. The wall of the flame cell has microvilli where it joins the next cell of the tubule, and the resulting slits function as a filter. The beating of the cilia creates a negative pressure and interstitial fluid is drawn into the tubule through these slits. Molluscs and most annelids possess metanephridia. The inner end is not blind but has a nephrostome funnel opening into the coelom. Earthworms possess one pair of such tubular organs per segment.
Green (antennal) glands are found in crustaceans such as crabs. The organ has a saccule in the head, bathed in hemolymph. The saccule has interdigitating podocyte cells allowing the filtration of fluid from the hemolymph (the cells are also found in protonephridia and in vertebrate kidneys). From the saccule, fluid passes to a labyrinthine tubule where selective resorption occurs; the resulting urine is stored temporarily in a bladder in some species before being expelled via a pore at the base of the antenna. (The saccule is a remnant of the coelom: thus the tubule may be homologous to a metanephridium.)
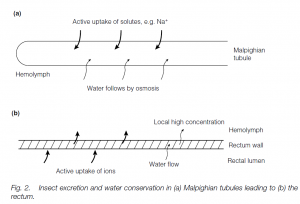
Insects and spiders possess Malpighian tubules, blind-ending tubules bathed in hemolymph (compare Crustacea). There are from two to several hundred tubules which open into the intestine. Filtration does not take place into the tubules: uric acid (as soluble potassium urate), potassium and sodium are secreted from the hemolymph into the tubules and water follows. The contents then pass down to the rectum. Ions are actively pumped out of the fluid and again water follows; uric acid crystallizes out. Thus hypertonic urine (particularly in species from arid habitats) is produced and water is conserved.
Vertebrate kidney
Most vertebrate kidneys work on the principle of ultrafiltration of plasma (leaving cells and macromolecules behind) from the high-pressure, closed blood system, with elements of selective solute and water resorption and secretion superimposed.
The filtration/resorption system can process large quantities of fluid, but often 99% of the fluid is resorbed: a secretory kidney would seem to imply less energy expenditure. Some teleost fishes have exclusively secretory kidneys; however, this does not allow them to eliminate novel toxins effectively in a changed environment, Ultrafiltration is nonselective with respect to small molecules. All vertebrate kidneys can produce urine hypotonic or isotonic with respect to the blood; only birds and mammals can make hypertonic urine.
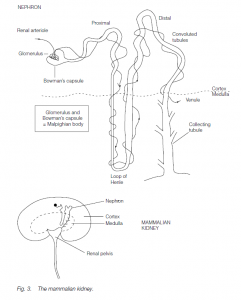
Each kidney is made up of several million nephron units comprising glomeruli and kidney tubules . Ultrafiltration (filtration powered by the hydrostatic pressure of arterial blood) occurs in the Malpighian corpuscle: a knot of capillaries (the glomerulus) is embedded in the hollow cup-shaped end of the kidney tubule, the Bowman’s capsule. The ultrafiltrate passes down a proximal convoluted tubule, round the hairpin bend of the loop of Henle, through the distal convoluted tubule to the collecting duct. The collecting ducts from all the tubules drain into the renal pelvis from which urine passes out via the the ureter, often being stored temporarily in the bladder.
Blood pressure forces fluid out from the thin walls of the glomerular capillaries: the hydrostatic pressure of the arterial blood must exceed the colloidal osmotic pressure of the plasma proteins (if the renal artery is clamped, ultrafiltration ceases). In amphibians (which have Malpighian bodies near the kidney surface), the ion, urea and glucose concentrations in the Bowman’s capsule are the same as in plasma. Inulin (a non-toxic polysaccharide that can be injected into the bloodstream) is not metabolized, resorbed or secreted, but is filtered: if urine inulin is at 100 times its plasma concentration, then 99% of water in the ultrafiltrate must have been resorbed. In a normal man, the filtration rate is more or less constant at approximately 130 ml per minute, 780 ml per hour, about twice the plasma volume; thus, resorption of water must be very efficient.
Glucose is actively resorbed in the proximal convoluted tubule: normal plasma glucose levels are about 100 mg per 100 ml; if this is exceeded the amount of glucose filtered and resorbed can rise to 230 mg per ml. This is the tubular maximum, TM. Above this level, plasma glucose spills over into the final urine, resulting in glucosurea, as in diabetes mellitus. Salts are also resorbed, and water follows (about 85% of filtrate water is resorbed here in humans).
Tubular secretion eliminates phenolics and their detoxification products, and penicillin and other xenobiotics. Phenol red excretion has been used to determine renal blood flow: it is removed completely by the kidney. Phenol red can be injected into the bloodstream, and the amount appearing in urine after a given time noted.
Adult man: renal plasma flow = 0.7 liters per minute
hematocrit = 45%
therefore blood flow = 1.25 liters per minute
(approximately one-quarter of the resting cardiac output)
Hypertonic urine in birds and mammals
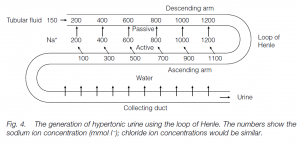
Birds can make urine with up to twice the plasma concentration of salts; mammals up to 25 times (desert mammals are most successful at this). The process is based on Na+ and Cl– resorption, using the loop of Henle: the concentrating ability is related to the length of the loop. Most mammals have nephrons with long and short loops; desert species (e.g. jerboa) and marine mammals (e.g. whales) have only long loops, fresh-water aquatic species (e.g. beaver) have mainly short loops.
Ultrafiltrate fluid enters the loop with a Na+ concentration, [Na+], equal to the plasma concentration (150 mmol l–1). Chloride is actively transported out of the ascending arm of the loop and there is also positive movement outwards of sodium ions; this increases the [Na+] in the surrounding tissue to, say, 200 m mol l–1. The descending arm is permeable to sodium and to water, and sodium diffuses in until the [Na+] in the descending arm is equal to that in the surrounding tissue (200 mmol l–1). The ascending arm must be water-impermeable to prevent water passively following the sodium ions into this arm. Fluid flowing out of the loop into the distal tubule is sodium depleted. Therefore the [Na+] is less than that in the plasma. As the fluid which leaves the loop has less sodium than that which entered, sodium must accumulate and concentrate in the loop in higher and higher concentrations: the longer the loop, the higher the concentration. This is a counter current multiplier system. The fluid entering the collecting tubule which parallels the loop is very dilute: the wall is water-permeable and water flows out by osmosis into the high [Na+] of the surrounding tissues. This leads to concentrated urine. Urea also plays a role in the concentration gradients. Birds are less effective at producing hypertonic urine: they excrete uric acid, and too concentrated a uric acid paste would clog the tubules. There is some evidence that they resorb water in the cloaca. Marine birds eliminate excess salt using nasal glands.
Regulation of urine
Human urine flow can vary from 10 to 1000 ml per hour. About 85% of water is withdrawn in the proximal tubules as a consequence of salt and water resorption. The distal convoluted and collecting tubules are responsible for the concentration of urine: the process is partly controlled by anti-diuretic hormone, ADH (vasopressin). ADH is secreted by the hypothalamus, passing via the posterior pituitary to the bloodstream. ADH acts by increasing the water permeability of the collecting ducts.
- When the body is overloaded with water, ADH secretion ceases and the collecting tubules become impermeable to water, leading to copious, dilute urine (diuresis).
- When the body is dehydrated, ADH is secreted making the collecting tubules more permeable to water, leading to small volumes of concentrated urine.
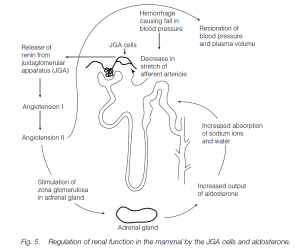
The osmotic concentration of blood is sensed by osmoreceptors in the hypothalamus: shrinkage of cells stimulates ADH secretion (and also a thirst urge). (ADH has a similar effect on frog skin, making the skin more permeable to water and stimulating osmotic inflow of water when desiccated.) In diabetes insipidus there is a pathological lack of ADH, resulting in chronic diuresis (copious urine flow) and excessive thirst. If the body is overloaded with water but low on salts ADH secretion ceases (low plasma osmolarity) and maximum recovery of sodium occurs in the distal tubule. This is stimulated by aldosterone secretion by the adrenal cortex: it accelerates the sodium pumps in the distal tubule. If aldosterone is accompanied by ADH secretion, water follows sodium isosmotically out of the tubule and into the blood. If ADH is absent (but aldosterone is present), the tubule is waterproof: sodium is extracted from the tubule leaving water behind. Thus aldosterone facilitates sodium removal and conservation. Aldosterone itself is stimulated by a decrease in blood pressure in afferent arterioles of the kidney (e.g. following hemorrhage) and decreased [Na+] in the early part of the distal tubule. When the blood pressure falls, juxtaglomerular apparatus (JGA) cells release renin. Renin acts on angiotensinogen (from the liver) to convert it to angiotensin I and this is later converted to an octopeptide angiotensin II. Angiotensin is a powerful vasoconstrictor, so blood pressure rises and glomerular filtration rates are restored. It also stimulates the synthesis and release of aldosterone (see above), and so sodium retention. Thus the kidney acts as a homeostat to maintain its own glomerular filtration rate. (Decreased sodium in distal tubules is detected by macula densa cells in the tubule wall, also leading to renin release by JGA cells, aldosterone and sodium retention. If [Na+] falls relative to [K+], aldosterone is released, and sodium pumps in the distal tubules exchange sodium in the filtrate for potassium in the plasma).
Evolution of the vertebrate kidney
Ancestral vertebrates probably possessed a segmental kidney, with one pair of nephrons for each body segment between the anterior and posterior ends of the coelom. They drained into an archinephric duct. Such a kidney is a holonephros.
In adult fishes and amphibians, the anterior tubules are lost, the middle tubules are associated with draining the testis in the male, and the functional kidney is the hind-most part of the holonephros: this is known as an opisthonephros. The archinephric duct functions in most amphibians and fishes as a ureter and a sperm duct (teleost fishes have a separate sperm duct). Reptiles, birds and mammals (amniotes) have a secondarily non segmental metanephros with its own ureter. Most of the old opisthonephros is now (in males) the epididymis associated with the testis, and the archinephric duct becomes the vas deferens.
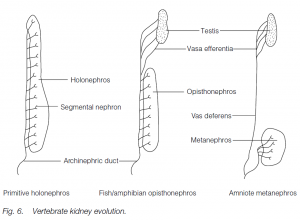
During amniote embyrogenesis, there is a process which partially parallels this evolutionary sequence: there are transitory pronephros and mesonephros in the front and middle portions of the embryonic nephrogenic mesoderm. The most posterior part develops to form a metanephric kidney.
References