Phylum Chordata
Classification: Kingdom Animalia, Sub-Kingdom Metazoa, Phylum Chordata, Common Names: sea-squirts, salps, lancelets, fishes, amphibians, reptiles, birds, mammals
Features
Type of organism
The Chordata is a very large and diverse phylum which has been studied extensively, mainly because it includes the vertebrates. Invertebrate chordates exist, however, and these are important in evolutionary terms. These are sometimes called protochordates, a term which formerly included the Hemichordata too. There are about 45 000 species of chordates, occupying marine, fresh and brackish water and terrestrial habitats. Several groups have evolved flight.
Body plan
Chordates are bilaterally symmetrical, triploblastic, segmented coelomates which demonstrate a deuterostomic pattern of early embryonic development. At some stage in their development they possess: a dorsal tubular nerve cord, formed from an infolding of a strip of dorsal ectoderm; a notochord; pharyngeal gill-slits; and, usually, a post-anal tail. Segmented muscles are found in the body.
Feeding
Invertebrate chordates are suspension-feeders: food particles are trapped in mucus on elaborately expanded pharyngeal gill-slit systems and wafted into the gut by cilia; modern jawless vertebrates are semi-parasites on jawed fishes; most jawed vertebrates are herbivorous or carnivorous, macrophagous feeders. Teeth inserted into vertebrate jaws are almost universal. The gut is complete with a mouth and an anus. An endostyle along the floor of the pharynx in invertebrate chordates is the homolog of the vertebrate thyroid gland.
Locomotion
Most chordates are active swimmers, using muscles which act against the antitelescopic notochord or vertebral column. (Many urochordate larvae metamorphose to form sessile adults.) The evolution of median and paired fins in fishes facilitates control of pitching, yawing and rolling. In land vertebrates, the paired fins have evolved to form jointed limbs which act as levers against the substratum: these may be secondarily lost (as in snakes). A number of chordate groups have evolved flight and others have resumed an aquatic habitat, usually using modified paired limbs.
Skeleton
Chordates are characterized by an endoskeleton; in the vertebrates the endoskeleton is made of cartilage, normally replaced by bone (consisting primarily of hydrated calcium phosphate and protein). The notochord forms as an antitelescopic, cartilaginous rod dorsal to the gut and ventral to the nerve cord. It contains a gelatinous matrix surrounded by tough connective tissue. In vertebrates, it is partly or completely replaced by vertebrae which develop intersegmentally and surround the nerve cord.
Respiration and vascular system
The pharyngeal gill-slits, used for suspension-feeding in invertebrate chordates, are used for respiration in the more advanced aquatic forms but are present only transiently in the embryo in land vertebrates. Gills develop in fishes and are present in amphibian larvae. Land vertebrates respire using lungs which develop as pouches from the gut (and are the homolog of the fish swimbladder). Most chordates have a high-pressure, closed circulation with a ventral heart. A dorsal vessel conveys blood backwards while a ventral vessel conveys it forwards.
Osmoregulation and excretion
The characteristic excretory organs of vertebrates are the segmental kidneys, replaced in reptiles, birds and mammals by nonsegmental kidneys; invertebrate chordates rely on simple diffusion of waste substances (urochordates) or groups of solenocytes (cephalochordates).
Co-ordination
The dorsal hollow nerve cord, present at some stage during the chordate’s life history, is characteristic of this phylum. In metamorphosed urochordates the nerve cord is lost. In vertebrates the anterior end of the nerve cord expands to form a brain.
Reproduction
Reproduction is normally sexual, although parthenogenesis is seen in a few species, and some urochordates reproduce asexually by budding to form colonies.
Sub-phyla
There are three chordate sub-phyla:
(1) Urochordata
(2) Cephalochordata
(3) Craniata (= Vertebrata)
The first two are sometimes called the acraniate or invertebrate chordates. The craniates are more popularly termed the vertebrates.
Sub-phylum Urochordata (Tunicata)
The 1300 species of urochordates or tunicates have larvae with the typical chordate structure (including a notochord, a dorsal nerve cord and a segmentally muscled tail). However, after metamorphosis this structure, apart from the gill slits, is lost. The tail with its musculature and the notochord disappear, while the nerve cord is reduced to a round ganglion from which peripheral nerves radiate. The pharynx enlarges to form a branchial basket with numerous ciliated gill-bars covered in mucus. Water is drawn through the mouth, passing through the gill-slits to an atrium. Food particles are filtered out in the mucus wafted by cilia along the gill-bars. They pass to a dorsal epipharyngeal groove and thence back to the gut.
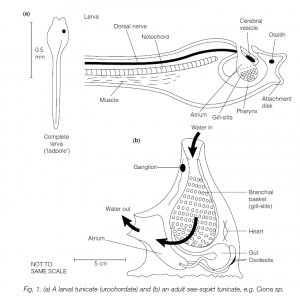
Ventrally in the pharynx is an endostyle, which concentrates iodine and is the homolog of the vertebrate thyroid gland. The body is enclosed in a cellulose ‘test’ or tunic. Urochordates have a heart (which can reverse its direction of beat) and blood vessels. Functional lymphocytes are seen, and rudiments of the vertebrate adaptive immune system have been described. Blood cells include vanadocytes (containing hemovanadium, although this vanadium-containing pigment has not been demonstrated to have a respiratory function). Urochordates are usually hermaphrodite with an ovotestis. Eggs develop into a ‘tadpole’ larva which metamorphoses into an adult. Many multiply asexually by budding to form a colony with a common exhalant opening from the atrium.
There are three classes: the Ascidacea, the Larvacea and the Thaliacea.
Class Ascidiacea: sea-squirts
Usually sessile and bottom-living. They may be solitary or colonial. Examples include Ciona intestinalis and Botryllus sp.
Class Larvacea
Forms with a neotenic retention of some larval features, such as a tail, into adulthood. Oikopleura has an elaborate, secreted ‘house’ used for filter-feeding. An example is Oikopleura dioica.
Class Thaliacea
Salps, solitary or colonial. The test has muscle bands and the atrial opening points posteriorly allowing the salp to swim by jet-propulsion. An example is Salpa sp.
Sub-phylum Cephalochordata
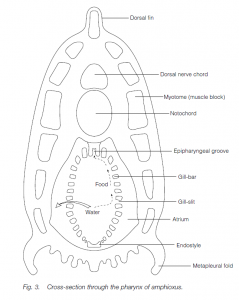
The Cephalochordata comprise two genera. In Branchiostoma lanceolatum (= Amphioxus lanceolatum), the lancelet or amphioxus , the notochord persists throughout life: it extends to the extreme anterior end, beyond the hollow nerve cord which lies dorsal to it; the nerve cord does not expand to form a brain and there is little cephalization. The animal lies half-buried in the sand, suspension-feeding particles from the sea water. Cephalochordates are small (<5 cm). A specialized feature of the sub-phylum is the asymmetrical body architecture whereby segmented muscles and nerves alternate on each side of the body. (The second cephalochordate genus is called Asymmetron.)
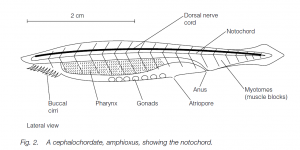
The pharynx is long with many ciliated gill-bars: the cilia generate a current so that water is drawn into the mouth, passes through the gill-slits where food is filtered out, and then enters the atrium and thence out through the atriopore. Mucus is produced by a ventral endostyle (the homolog of the vertebrate thyroid gland) in the pharynx. Similarities with the feeding patterns of urochordates are seen. Gonads are arranged segmentally: the sexes are separate. Excretion is facilitated by groups of flame cells (solenocytes) similar to those in platyhelminths and polychaete annelids. The blood vessel flow pattern is similar to that in vertebrates, but there are no blood cells nor a heart: the vessel walls are contractile. An example is Branchiostoma lanceolatum (amphioxus or lancelet).
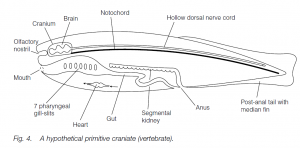
Sub-phylum Craniata (Vertebrata)
Craniates possess the diagnostic chordate features (at some stage in their life histories) of a notochord, hollow dorsal nerve cord, pharyngeal gill-slits and a post-anal tail. All craniates have an anterior expansion of the dorsal nerve cord, the brain, contained in a cranium (the brain-box or skull). This cephalization is further marked by extensive development of sense organs in the head. The notochord is usually replaced in part or totally by intersegmental cartilage or bony units, the vertebrae. Cartilage is found in some invertebrates such as Mollusca, but bone is unique to vertebrates, mainly consisting of hydrated calcium phosphate minerals in a protein matrix. There is a ventral heart behind the head. Blood flows forward ventrally from the heart, then through the gills, and then upwards and backwards dorsally. The excretory organs, the kidneys, lie dorsally in the coelom: they comprise thousands of nephron units. The sub-phylum is normally divided into two superclasses: the agnathans and the gnathostomes.
Superclass Agnatha (jawless vertebrates)
The earliest known vertebrates, the ostracoderms, lacked jaws: these armored fishes are totally extinct. Modern Agnatha belong to the class Cyclostomata (round mouths): this comprises the lampreys and the hagfishes . The absence of jaws is diagnostic. Cyclostomes have round, suctorial mouths. They are semi-parasites on larger, jawed fishes to which they attach, rasping at the host flesh with a toothed tongue and horny teeth within the mouth cavity. The sexes are separate. The fertilized eggs of lampreys hatch to form filter-feeding ammocoetes larvae which lie in the mud; after some years they metamorphose into lampreys.
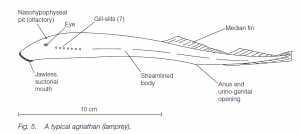
Specialized features of lampreys include the mouth and tongue, the streamlined shape, and pouched gills allowing a tidal flow of water over them.
Primitive features include a persistent notochord, a lack of paired fins, no ducts from the gonads and seven pairs of gills. The skeleton is cartilaginous and includes a cranium, supports anterior to each gill pouch (gill arches) and peg-like vertebrae.
Degenerate features include reduced eyes and a simple, straight gut. Examples include Lampetra fluviatilis (common lamprey) and Myxine glutinosa (Atlantic hag).
Superclass Gnathostomata (jawed vertebrates)
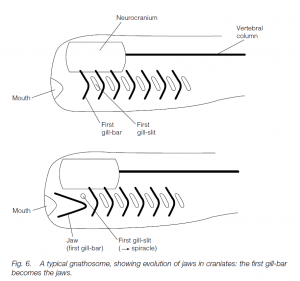
The Gnathostomata includes all other vertebrates. In gnathostomes the first gillbar becomes ‘wrapped round’ the mouth to form the upper and lower jaws. This allows feeding on large particles (macrophagy), especially when teeth are inserted into the jaws. Eight classes of gnathostome are usually recognized.
(1) Class Acanthodii. The classification of the extinct acanthodians, known from the Devonian, is problematical: they exhibit affinities to cartilaginous fishes, to bony fishes and to forms unknown today. They are the first fishes to have true paired fins (unlike the paddles of some ostracoder magnathans) and to possess jaws. The paired fins were attached to the trunk via limb girdles, and there were intermediate, paired spines, between the pectoral and pelvic fins, apparently not associated with fin structures. An example is Climatius sp.
(2) Class Placodermi. The placoderms were Palaeozoic shark-like fishes with jaws and an armor of bony plates. There were several subclasses, now all extinct. An example is Dunkleosteus sp.
(3) Class Chondrichthyes. First known from the upper Devonian, the Chondrichthyes (cartilaginous fishes) have two subclasses. With the exception of only four or five species they are marine. Chondrichthyes have a cartilaginous skeleton (which may be secondarily calcified) and scales, identical in structure to teeth (including dentine). Pectoral and pelvic paired fins are present. Fertilization is internal, with the females laying large, yolky eggs or exhibiting (ovo)viviparity.
(a) Subclass Elasmobranchii.
The elasmobranchs include the sharks, dogfishes, skates and rays. The upper jaws move with respect to the cranium; there are 5–7 gill-slit pairs and an anterior spiracle, each opening separately to the exterior. The order Selachii includes the sharks, many of which are large predators; the order Batoidea includes skates and rays with dorso-ventral flattening and enlarged pectoral fins. Many batoideans are molluscivores with flattened, crushing teeth. Examples include Carcharodon carcharias (great white shark); Manta sp. (manta ray).
(b) Subclass Holocephali.
The deep-sea rat-fishes, rabbit-fishes and chimeras may not be closely related to the sharks and rays. Their skeletons, gill and vascular systems, and their nervous systems are different from those of the Elasmobranchii. An example is Hydrolagua colliei (rat-fish).
(4) Class Osteichthyes. With more than 30 000 species, the Osteichthyes (bony
fishes) include more species than all the other vertebrate classes put together. The embryonic cartilage in the skeleton is replaced by bone. Paired fins are attached to limb girdles. The gill-slits open to an opercular cavity which is enclosed within an operculum. Pharyngeal pouches from the foregut were probably primitive lungs: these may be modified to form a swim-bladder, used for buoyancy. Fertilization of the eggs is external: large numbers of small eggs develop into larvae (fry) which metamorphose into adults.
There are two subclasses:
(a) Subclass Actinopterygii.
The Actinopterygii or ray-finned fishes have bones and muscles restricted to the bases of the fins which are supported distally by fin rays. The bony scales have a thick layer of enamel. Almost all bony fishes alive today are actinopterygians: three grades or infraclasses are recognized, with progressively more advanced features:
- All early actinopterygian fossils belong to a group of Chondrostei known as the Palaeonisciformes (palaeoniscids) which radiated extensively between Carboniferous and Triassic times; the modern Chondrostei are relics and include the bichir, the paddle-fishes and sturgeons. Air-sacs or lungs are present. Examples are Polypterus ornatipinnis (bichir) and Huso huso (Volga sturgeon).
- At this grade, morphological body changes allow for more efficient swimming and feeding. Most Holostei died out in the Cretaceous and the Eocene, but survivors include the bowfin and the gar-pike. Examples are Amia calva (bowfin); Lepisosteus osseus (gar).
- Most living fishes are teleosts (about 30 000 species): the grade or infraclass radiated extensively during the early Tertiary. Teleosts have thin, bony scales without enamel, a moveable upper jaw which hinges anteriorly to the skull, a complete vertebral column which replaces the notochord and a single, dorsal swimbladder which may or may not be connected to the gut. Teleost classification is complex: many zoologists feel that they show polyphyletic (multiple evolutionary) origins from the Holostei. Examples are Cyprinus carpio (carp); Esox lucius (pike).
(b) Subclass Sarcopterygii.
The sarcopterygians or fleshy-finned fishes have paired fins with an axis of bone and muscle. There are two intra classes:
- The main importance of the crossopterygians (the tassel-finned fishes) lies in the fact that from them probably arose the land vertebrates (the Tetrapoda). Forms such as Osteolepis sp. have similar limb bone and skull bone arrangements to the earliest amphibians. There is only one living species, the coelacanth. This ovoviviparous fish (from deep waters off the Comoro Islands in the western Indian Ocean and elsewhere) has a primitive blood system and a fat-filled swim-bladder. The living example is Latimeria chalumnae (coelacanth).
- The true lungfishes survive today in three living species, from Australia, South America and East Africa respectively, being relics of a group abundant in Devonian fresh waters. Dipnoans have lungs and breathe air but they also retain their gills. The African and South American forms are obligate air-breathers, but the Australian lungfish needs oxygenated water to survive. The blood system is, for fishes, sophisticated, with a partially divided heart and with pulmonary veins returning oxygenated blood to the left atrium. Estivation in cocoons the mud is common during drought periods. Male African and South American lungfishes brood eggs which hatch into larvae with external gills. Examples include Neoceratodus forsteri (Australian lungfish); and Lepidosiren paradoxa (South American lungfish).
(5) Class Amphibia. Amphibians arose from the crossopterygian fishes during the early Devonian: these early amphibians were known as Labyrinthodontia on account of the architecture of their teeth. Modern forms, collectively known as Lissamphibia, have problematical relationships with the labyrinthodonts.
Amphibians typically possess two pairs of legs (not fins), although one or both pairs may be secondarily absent. The pectoral girdle does not connect to the back of the skull; there is a pelvic girdle connected to the vertebral column by sacral vertebrae. The middle ear has a single bone connecting the eardrum to the inner ear.
Amphibian larvae (tadpoles) have external gills which may later be internalized. They develop in water, which may include specialized habitats such as pools in the bases of flowers or a brood pouch in the mother’s mouth. Metamorphosis is under the control of the hypothalamus– pituitary–thyroid axis and a prolactin–thyroxin balance. Metamorphosis may be profound: lungs replace gills (although many species also employ cutaneous respiration). Some urodele species become sexually mature without undergoing metamorphosis, exhibiting neoteny (retention of larval characters in the adult) and pedomorphosis (sexual maturity in the larva). Amphibian hearts have two atria and one ventricle.
There are three living orders of Lissamphibia:
(a) Order Anura.
The anurans are the toads and frogs which in the adult form lack tails. Sexual maturity is always preceded by metamorphosis from a tadpole larva. Examples are Rana temporaria (common frog); Rana catesbeiana (bullfrog); Xenopus laevis (African clawed toad).
(b) Order Urodela.
The newts and salamanders and their allies have long tails used for swimming. Forms range from those that are totally terrestrial, through facultative neotenes which are sexually mature as larvae and only metamorphose in response to drought or thyroxine treatment, through to perennibranchiates which are never observed to metamorphose. These forms retain their gills. Examples are Triturus cristatus (crested newt); Ambystoma mexicanum (Mexican axolotl).
(c) Order Apoda.
The apodans or gymnophionids are limbless, burrowing amphibians from the tropics or southern temperate regions; the larval stage may pass within the egg. An example is Dermophis mexicanus.
(6) Class Reptilia.
Reptiles evolved from labyrinthodont amphibians during the late Devonian. The ancestral group were the Cotylosauria or ‘stemreptiles’. From this group evolved various taxa including the Chelonia (tortoises and turtles), the Lepidosauria (the tuatara, lizards and snakes), the Synapsida (whose later members included the mammal-like reptiles which gave rise to the mammals), the Archosauria (whose living members include the crocodiles but which once claimed the flying pterosaurs and the Ornithischia and the Saurischia, the two ‘dinosaur’ orders) and several extinct groups of marine reptiles (such as ichthyosaurs and plesiosaurs). Like birds and mammals, reptiles have an amniote egg : the embryo develops an amnion, chorion and allantois in addition to a yolk-sac.
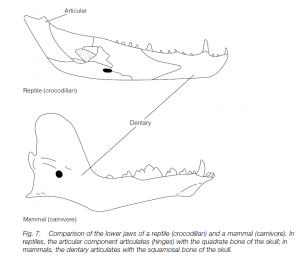
The three classes are sometimes grouped as a superclass Amniota. There are no larval stages, and some reptiles (e.g. sea-serpents and adders) are viviparous. The chorioallantois acts as a respiratory organ in the developing, waterproof egg. Gills are absent. Oviparous marine and freshwater species (e.g. turtles and crocodiles) must lay eggs on land. In reptiles the articular bone of the lower jaw articulates with the quadrate bone of the skull . Reptiles have a more-or-less waterproof skin, usually with scales. The kidney develops non segmentally and is known as metanephric (as opposed to the segmental, meso- or opisthonephric kidneys of fishes and amphibians). Reptiles reached their peak during the Mesozoic period. Modern forms belong to four orders, distinguished largely by the architecture of the skull.
(a) Order Rhynchocephalia.
This is a primitive group represented today solely by the rare ‘living fossil’ Sphenodon punctatus, the tuatara from New Zealand. It has primitive skeletal features and may resemble forms ancestral to the lizards.
(b) Order Squamata.
The familiar lizards and snakes have scales (and often plates too). The skull architecture shows characteristic fenestrations for each suborder. Members of the suborder Sauria (lizards) usually have four legs although some lizards (e.g. the slow-worm Anguis fragilis), the ‘wormlizards’ (suborder Amphisbaenia) and the snakes (suborder Serpentes) have lost their limbs and limb girdles (vestigial limbs are occasionally seen in pythons). The skull may be specialized to allow the swallowing of large prey, and some species are venomous. Snakes lack eardrums. Examples include Lacerta vivipara (common lizard); Vipera beris (adder).
(c) Order Chelonia.
In turtles and tortoises, the body is enclosed in a shell of bony plates fused to the ribs and the vertebrae. Limb girdles lie within the shell. A horny beak replaces teeth. The organ systems of chelonians are relatively primitive. An example is Chelonia mydas (green sea turtle).
(d) Order Crocodilia.
Crocodiles, alligators and caimans are the only living survivors of the Archosauria (ruling reptiles) which once included the ‘dinosaurs’, pterosaurs and other extinct reptiles of the Mesozoic period. The skin of crocodilians is covered with bony plates with horny scales outside. The lungs are more complex than the simple sacs seen in other reptiles, and the ventricle of the heart is almost completely divided to make an effective four-chambered heart. The skull architecture resembles that of the ‘dinosaur’ groups and birds. An example is Alligator mississippiensis (Mississippi alligator).
(7) Class Aves. In evolutionary terms, the birds are a very homogeneous class, probably derived from saurischian archosaurs (lizard-hipped ‘dinosaurs’) of the Jurassic. The skeletons of the earliest birds (e.g. Archaeopteryx lithographica) show remarkable similarities to coelurosaur ‘dinosaurs’ such as Deinonychus sp. The reptilian scales have evolved into feathers in birds. The forelimb is modified to form a wing, bearing primary feathers on the ‘hand’ for propulsion, secondary feathers on the forearm for lift and contour feathers for aerodynamic streamlining.
Down feathers provide insulation to maintain the homoiothermy (‘warm-bloodedness’) of this class. The skull has a large cranial vault to contain the big brain. The eye orbits are large (birds use vision as their main sense). There are no teeth in modern birds but there is a horny beak whose form reflects the diet of the species. The lungs are large and efficient; an associated air-sac system permits a unidirectional air-flow in the lungs, with blood vessels arranged in a crosscurrent fashion to permit extraction of up to 90% of the oxygen from inspired air. The heart has four chambers: there are separate, parallel systemic and pulmonary circulations. All birds are oviparous: eggs are laid on land, usually in a nest. Parental care of the young is normal.
There are two modern superorders:
(a) Superorder Palaeognathae.
The ratites are usually flightless. The palate is specialized. There is considerable debate over whether ratites are ancestral and/or more primitive than other living birds. Examples include Struthio camelus (ostrich); Apteryx haastii (kiwi).
(b) Superorder Neognathae.
This superorder includes all other birds. There are about 20 orders, the largest being the Passeriformes (perching birds, e.g. the robin). Examples include Gallus domesticus [domestic fowl (chicken)]; and Corvus corone (carrion crow).
(8) Class Mammalia
The mammals are characterized by the possession of hair (fur): scales (e.g. on a rat’s tail) are rare. The air trapped within the fur provides insulation to help to maintain the homoiothermy of this class. The only bone in the lower jaw is the dentary: this articulates with the squamosal bone of the skull; the redundant articulating bones of the reptile are used as auditory ossicles of the middle ear (the maleus and the incus, together with the stapes, hammer, anvil and stirrup).
Unless secondarily lost, teeth are present in the upper and the lower jaws: these show considerable diversity of form, adapted to diet. The young of mammals are suckled on milk secreted by the mother’s mammary glands. The lungs have alveoli. A tidal flow of air into and out of the lungs is assisted by a muscular diaphragm which separates the thoracic and abdominal cavities. The heart has four chambers: as in birds there are separate pulmonary and systemic circulations. Mammals evolved in the Triassic from synapsid reptiles. There is a well documented fossil series which leads from mammal-like reptiles (e.g. cynodonts and ictidosaurs) through to the true mammals. Fossil Mesozoic mammals are rare and are usually known from only jaws and teeth.
There are two subclasses:
(a) Subclass Prototheria (= Monotremata).
This subclass, with uncertain affinity to both fossil forms and to other modern mammals, lays eggs and the milk is produced by glands rather different from other mammals’ mammary glands. The skeleton is distinctive. There are three living species, the duck-billed platypus and the spiny anteaters, all from Australasia. An example is Tachyglossus aculeatus (short-nosed echidna).
(b) Subclass Theria.
The Theria (‘beasts’) includes all other living mammals. There are two infraclasses:
- Infraclass Metatheria (= Marsupialia). In marsupials, the young are born very immature; they crawl through the mother’s fur to the mother’s abdomen where they attach to a nipple, usually in a pouch. Most marsupials are native to Australia where the infraclass has radiated widely, but the opossums are indigenous to the Americas. Marsupials have epipubic bones on the pelvic girdle. An example is Setonix brachyurus (quokka).
- Infraclass Eutheria (= Placentalia). In placentals, the young grow within the uterus, using a chorioallantoic placenta. The degree of maturity at birth varies (cf. newborn mice and guinea-pigs). The infraclass has radiated widely. About 15 living orders are recognized, including:
- Insectivora
A primitive, arguably ancestral order retaining a vestigial cloaca and having a rather lower body temperature than other placentals. An example is Sorex araneus (common shrew).
- Chiroptera (bats)
Forelimbs are extensively modified to form a wing of skin. An example is Pipistrellus pipistrellus (common pipistrelle).
- Primates
Large eyes and brains, and with prehensile hands and (often) feet. Opposable thumb and the claws replaced by nails. Otherwise rather unspecialized: includes prosimians (e.g. lemurs and bushbabies), old and new world monkeys, apes and humans. Examples include Nycticebus coucang (slow loris); Macaca mulatta (rhesus monkey); Pan troglodytes (chimpanzee).
- Carnivora
Normally flesh-eaters, usually with four clawed digits on each limb. Examples include Panthera leo (lion); Odobenus rosmarus (walrus).
- Cetacea
Whales and dolphins are wholly aquatic; the hind limbs are lost. Some have teeth, others filter plankton with baleen plates on the palate. An example is Tursiops truncatus (bottle-nosed dophin).
- Proboscidea
The elephants have reduced numbers of teeth; the upper incisors are modified to form tusks, and the nose and upper lip are extended to form a trunk. An example is Loxodonta africana (African elephant). [Arguably related orders include the Hyracoidea (the hyraxes) and the Sirenia (the dugongs and manatees: marine herbivores.]
- Artiodactyla
The even-toed ungulates, usually with two hoofed digits on each limb. They are omnivores or herbivores, including ruminants with compound stomachs adapted to the symbiotic digestion of cellulose. Horns are common. Examples include Cervus elaphus (red deer), and Camelus dromedarius (Arabian camel).
- Perissodactyla
The odd-toed ungulates are herbivores with an odd number of hoofed digits on each limb, e.g. one in horses. Examples include Equus berchelli (zebra); Diceros bicornis (African black rhinoceros).
- Rodentia
The rodents are a very large and successful order (>3000 species). They have a single pair of upper and lower incisors for gnawing: the teeth grow continuously. There is a wide gap (diastema) between the incisors and grinding molars. An example is Rattus rattus (black rat).
- Lagomorpha
The rabbits, hares and pikas are similar to the rodents but possess two pairs of upper incisors. An example is Oryctolagus cuniculus (European rabbit). There are also three orders of edentates (armadilloes, sloths, anteaters, pangolins and the aardvark) which tend to eat ants, termites and other insects: they are characterized by long snouts, long, adhesive tongues and reduced or absent teeth.