Thermoconformers and Thermoregulators
Most animals are thermoconformers (whose body temperature conforms to that of the surrounding environment) but some animals are thermoregulators, either behaviorally or physiologically.
Body temperature in mammals and Birds
Heat produced by an animal must be transported to the surface before it can be transferred to the environment. Thus the surface is at a lower temperature than the core or there would be no transfer: the animal is not at a uniform temperature.
In humans, organs of the thorax and the abdomen, together with the brain, generate 72% of the body’s heat at rest. Core and shell temperatures are recognized. During exercise, the amount of heat to be transported to the body surface must increase by tenfold or more. Even in the core, the temperature is not uniform: some organs generate more heat than others, those near veins are cooler than those near arteries, etc. Deep rectal temperature is often used as a representative core temperature.
Diurnal temperature fluctuations occur, of about 1–2°C: the peak temperature is when the animal is most active (e.g. at night for noctural species). Such circadian (daily) rhythms are independent of exercise, occurring even in resting animals, and seem to be related to the photoperiod: the cycle is reversible by reversing the illumination cycle [i.e. a noctural species kept in a reversed light cycle will exhibit a peak temperature during the day, when it is artificially ‘night’ (darkness)].
Core temperatures:
● Monotremata (Prototheria) 31 ± 2°C
● Marsupialia (Metatheria) 36 ± 2°C
● Placentalia (Eutheria) 38 ± 2°C
● Aves 40 ± 2°C
‘Primitive’ orders of placentals (e.g. Insectivora) tend to have a lower core temperatures than do ‘advanced’ orders (e.g. Carnivora). Control seems to be sophisticated: related species of mammals from polar and tropical regions have very similar core temperatures; the same applies in birds. The core temperature of sheep, recorded using a telemetric thermocouple, is the same (± 0.1°C) whatever the weather at a given time of day, and only varies by ± 1.9°C during a whole year.
Regulation of body temperature is mainly under autonomic (parasympathetic) nervous control, although sympathetic fibers control vasodilation, and shivering is under somatic nervous control: temperature detectors are more useful near the peripheries of the body where most variation occurs. Warm neurons linked to sensors in the skin increase activity as the temperature rises whereas cold neurons increase activity as it falls. Information is also fed in from core temperature sensors. [Gain is a useful concept: it measures how far the temperature would shift without negative feedback divided by how far it does shift. For human thermoregulation, gain = 33 (i.e. temperature stress which would raise a human corpse from 37–47°C would actually raise a living, thermoregulating human by 0.3°C: gain = 10 ÷ 0.3 = 33).]
Heat transfer
If an animal is to maintain a constant temperature, heat loss must equal heat gain. Thus if the heat produced in exercise is 10 times that produced at rest, heat loss must also increase tenfold. Heat loss will relate to air or water temperature, wind or water currents, insulation, etc. Heat flows from higher to lower temperatures through the following mechanisms.
Conduction (direct transfer of kinetic energy of molecular motion between physical bodies in contact with each other); the thermal conductivity coefficient, k, measures the ease of heat flow:
k (water) = 5.8 × 10–3 joules sec–1 cm–1 °C–1
k (air) = 2.1 × 10–6 joules sec–1 cm–1 °C–1
k (fur) = 3.7 × 10–6 joules sec–1 cm–1 °C–1
Thus (dry) fur is a poorer conductor (and better insulator) than water. However, these values are complicated by curved surfaces. Convection is an extension of conduction and involves mass flow of molecules in a fluid; it may be free (due to density changes, warm air rises) or forced (e.g. fanning).
Radiation involves no direct contact between the heat source and its ‘sink’; for physiological purposes, air is considered transparent to radiation. The intensity of radiation is proportional to [temperature (in kelvins) of radiating surface]4; thus emission rises rapidly with temperature. At physiological temperatures, most objects (e.g. skin) emit in the middle infra-red. The emissivity equals the absorptivity if an object is in equilibrium with its environment; thus skin, fur, etc. have high absorptivities in the middle infra-red.
[N.B. heat loss by radiation from pigmented and unpigmented bodies is about the same: colour may be important for heat absorption from the sun (e.g. in basking lizards) which has its peak intensity in the visible range.]
Heat loss to cool surroundings = Q.
Q = C(T2 – T1)
where T2 – T1 = temperature difference and C = a combined constant reflecting insulation, etc.
Q = C(TB – TA)
where TB = body temperature and TA = ambient (environmental) temperature. Evaporation also uses much heat (through latent heat of evaporation of water ≈ 2400 joules g–1); thus water evaporating from a surface (e.g. skin, tongue) results in loss of a great deal of heat.
Heat balance
For homoiothermy: heat loss = heat gain.
Htotal = ± Hc ± Hr ± He ± Hs
where Total is total heat production (positive), Hc is conductive and convective heat loss/gain (positive for net loss), Hr is net radiation heat exchange (positive for net loss), He is evaporative heat loss (positive for net loss) and Hs is heat stored (positive if the heat is stored and the body gains heat; temperature rises).
Values to the right of the equation can be negative (e.g. He can be negative when a cool body enters a hot, damp atmosphere, e.g. a Turkish bath). In a heat gain situation, a constant temperature can be maintained if evaporative heat loss increases.
Keeping warm
H = Q = C(TB – TA)
where H is heat production and Q is heat loss. The animal must tolerate TA(the environmental temperature) unless it moves; therefore it must adjust C (conductance) or TB(body temperature). Normally body temperature stays constant, so heat production or conductance must be altered.
● more exercise (more metabolic heat).
● involuntary muscle contraction (shivering).
● non-shivering thermogenesis [fat in brown adipose tissue (BAT) can be oxidized: it contains a protein which uncouples oxidative phosphorylation from adenosine triphosphate (ATP) production in mitochondria so that energy from oxidation is released as heat].
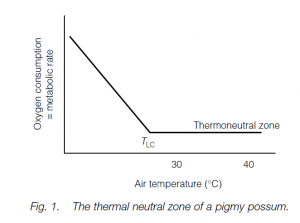
The metabolic rate stays constant as the air temperature falls and (TB – TA) increases: this is the thermoneutral zone. Below the lower critical temperature (TLC) the metabolic rate rises. TLC has lower values in well-insulated, polar species, and the gradients of the slopes showing the increase in metabolic rate with decreasing temperature are steeper in poorly insulated species.
Conductance measures heat flow from an animal to its surroundings. Insulation is the reciprocal of conductance; effective insulation minimizes heat loss. Heat loss is also reduced if the surface area: volume ratio is smaller. Larger animals conserve heat more effectively than do small ones with similar body architectures, and rolling into a ball or huddling can effectively reduce surface areas.
Fur is the major barrier to heat flow in land mammals: its insulation is very variable. Insulation increases with fur thickness, reaching a maximum in animals such as arctic foxes. Smaller animals have shorter, lighter fur and tend to be fossorial (burrowers) or hibernators. Polar bears have open, coarse fur which poorly insulates and, in water, most insulation is lost (loss in agitated water is 50 times that in air). In many species, winter fur is much heavier than summer fur: a black bear’s summer fur has only 52% of the insulation capacity of winter fur.
Many marine mammals and birds (e.g. seals and penguins) inhabit polar seas. Water has a high conductance. In the H = C(TB – TA) equation, C or H is altered. The metabolic rate in some marine species is higher than expected: in a harp seal, the metabolic rate is constant down to the freezing point of water. Insulation is by subcutaneous blubber (fat).
A countercurrent heat exchanger is found in flippers which lack blubber: returning venous blood is warmed by outgoing arterial blood. Each artery is surrounded by many veins; summer overheating is avoided by a venous shunt (the artery expands and compresses the veins) so that returning blood passes along alternative superficial veins and is not warmed by the arterial blood.
Such countercurrent heat exchangers are used in terrestrial homoiotherms: they allow a heron to stand in icy water, and are also found in human arms and legs Birds, with insulating down feathers, do not compensate completely by increasing metabolic rate with declining temperatures, that is in the H = C(TB – TA) equation, C (conductance) may drop with falling temperature. Other strategies include raising feathers and drawing the feet up to assume a ball-shape, allowing the shell temperature to drop while the core remains the same.

Keeping cool
Above the lower critical temperature TLC, metabolic heat production stays about constant throughout the thermoneutral zone. In the H = C(TB – TA) equation, if H and TB stay the same, then if TA rises, C must alter.
Heat flow is helped by vasodilation of cutaneous blood vessels and by increasing the surface area. As the environmental temperature goes up, the conditions for losing heat by conduction, convection and radiation become less favourable; evaporative heat loss and heat storage are more emphasized.
For example, a human in a desert has a metabolic rate of 293 kJ h–1 (≡ 0.12 litres of water evaporation); in the desert, a human may sweat 1–1.5 litres per hour (e.g. 1.32 l h–1). If there is no change in TB, then 1.2 litres of water (1.32 – 0.12) evaporated must be equivalent to the heat gain from the environment: thus the heat load from the environment is 10 times that from metabolism.
If TB can be raised by 1°C, then heat stored is 250 kJ, saving 0.1 litres of evaporated water. Surface area: volume ratio is again important, as heat gain from the environment is a surface process. Environmental heat load is related to the surface area while metabolic heat production is related to volume.
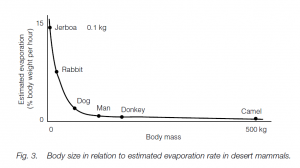
Thus, in arid conditions such as hot deserts, a small animal is at a disadvantage with respect to heat loads. It is possible to predict how much water animals will need to evaporate in order to dissipate heat: a jerboa will need to evaporate 14% of its body weight per hour, a rabbit 7% and a camel 1%; that is an exponential function of body weight.
Thus small animals tend to be fossorial (burrowing) and/or nocturnal (e.g. jerboa); the desert jackrabbit shelters in shadows during the day and vasodilates the extensive veins in its ears to lose heat. The camel derives advantages from its large size (and low area: volume ratio). If it thermoregulated in the same way as humans do it would lose about half of the percentage of water per hour, relative to body weight, that a human would.
However, it also increases Hs. A dehydrated camel has great diurnal TB fluctuations, from 34 to 41°C. For a 500 kg camel, a 7°C TB rise ≡ 12.1 MJ ≡ 5 litres of water At night the camel loses heat by radiation; (TB – TA) is also reduced as the camel heats up, and this saves water because less is used to defend TB by evaporative cooling. The camel is also helped by light-coloured, insulating fur [a shaved camel (!) would evaporate 50% more water]. Camels also tolerate more dehydration (10–12% body water loss is fatal in humans; 24% body water loss is tolerated by camels).
Ground squirrels store heat: they dart around and heat up quickly (large area: volume ratio) but cool off quickly in their burrows. Sweating and panting are methods used to evaporate water and lose latent heat of evaporation:
● Sweating is useful for naked mammals (e.g. humans) or in very dry atmospheres (e.g. camels); sweating is more passive than panting but results in a salt loss.
● Ungulates (hoofed mammals) and carnivores tend to pant. Many desert animals have long respiratory tracts which act as heat exchangers and water conservation channels – ingoing air is warmed and moistened in the nasal tract, outgoing air is cooled and water condenses. When cooling is required, the outgoing air is diverted over the moist buccal (mouth) membranes from which water is evaporated. Panting can lead to respiratory alkalosis and requires work – therefore, use is made of shallower respiration at greater frequency, using dead space in the upper respiratory tract – a dog panting shifts from 30 to 40 breaths per minute to 300 to 400 breaths per minute, the natural oscillation frequency of the respiratory system. Panting is best under high and sudden heat stress (e.g. a pursued gazelle): the core temperature gets very high but the brain stays cool, the carotid artery passing around a venous sinus near the nasal cavities.
● Rats salivate over their limbs in thermal emergencies.
● Pigs and hippopotami wallow in mud: the damp mud clings to the body and evaporates slowly – this technique uses external water.
Thermoregulation in poikilotherms
In most poikilotherms TA = TB. Small aquatic animals lose heat rapidly because of the high conductivity of water, and they cannot have too high a metabolic rate because of oxygen demands – huge gills would be required and these would be sources of heat loss unless a heat-exchanger was built in.
Many large, active fishes (e.g. bluefin tuna) isolate their swimming muscles and use a countercurrent heat exchanger to conserve metabolic heat, that is they are partial endotherms. Cutaneous arteries supply the swimming muscles. Parallel fine arteries enter the muscle, enmeshed in parallel veins leading out to the cutaneous vein; veins warm arterial blood as it enters.
The interior of the muscle can be more than 14°C above the water temperature. High temperatures increase muscle power output, enabling them to contract faster. Other heat exchangers isolate the tuna heart, liver, etc. On land, radiation and evaporative heat loss and gain are important.
To raise the temperature it is important to maximize gain from radiation and storage and minimize conductive and convective loss. Many lizards bask in the sun, gaining radiant solar heat and absorbing conductive heat from hot rocks. Liolemus sp., in the morning sun at –2°C, raises its cloacal temperature to 33°C within 60 minutes (at an ambient temperature of 1.5°C). This preferred (eccritic) the temperature of 33°C is maintained by moving in and out of the sun, orienting the body parallel or at 90° to the sun’s rays, and lifting the body (or two legs) off the rocks.
The phenomenon is known as heliothermy and is an example of behavioral thermoregulation. Some species alter the colour of their skin, making it darker in order to absorb more incident solar radiation in the visible wavelengths. In the marine iguana, cooling when swimming in a cold sea is much slower than heating, due to changes in skin circulation.
The lizard Sauromalus obesus pants when hot. Because the carotid arteries pass near the pharynx wall panting is adequate to cool the brain, but not the whole body. Brooding pythons ‘shiver’ to generate metabolic heat to warm their eggs.
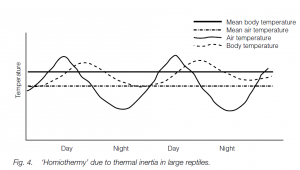
Thermal inertia may maintain large reptiles (e.g. monitor lizards, and perhaps applied in Mesozoic ‘dinosaurs’) at a more or less constant body temperature. The small area:volume ratio means that such animals heat up only a little by day and cool only a little by night, and metabolic heat may not escape.
Some flying insects warm their flight muscles by shivering. The sphinx moth Manduca sexta flies on cool nights with muscles at 35°C. The moth shivers and,the wings lightly vibrate due to spontaneous contraction of upstroke and downstroke,muscles. Thoracic heat is retained by furry scales; overheating is prevented by circulating hemolymph through the abdomen.
Torpor and Hibernation
Keeping the core temperature (TB) high is expensive, especially in small animals, with high metabolic rates. Therefore TB could be allowed to drop. Many small, mammals (e.g. dormouse, hedgehog) and a few birds (e.g. poorwill) hibernate over winter; other small animals undergo periods of diurnal torpor (e.g. bats,, hummingbirds).
During hibernation/torpor the metabolic rate falls and the, animal becomes torpid and less responsive to external stimuli. (Torpor is, confined to small homoiotherms: bears have a deep winter sleep but TB is little, depressed.) Estivation is a period of summer or drought inactivity.
For example, Columbian ground squirrels disappear into burrows from August to May., Some snails and lungfishes radically reduce their metabolic activities during, droughts., Torpor is characterized by lowered metabolic rates (oxygen consumption can be 2.5% of normal) and extreme bradycardia, including periods of asystole,(cessation of heartbeat): this can lead to acidosis.
Hypoxia is tolerated because, of high hemoglobin levels and a leftward shift of the oxygen–hemoglobin dissociation, curve. Endocrine function is quiescent, although gonads may become, active towards the end of hibernation., Hibernators, feed before hibernation. The phenomenon is triggered by the, time of the year, related to endocrine cycles partly supplemented by environmental, cues such as photoperiod, temperature and/or availability of food supplies.
Hibernation and torpor are under close control: the analogy is with turning down the thermostat in a heating system. The thermostat is the hypothalamus: thermoreceptors input to the hypothalamus and appropriate heat-generating or heat-loss mechanisms are elicited. Sensors can be bypassed experimentally by locally heating or cooling the hypothalamus using Thermo probes: the temperature at which no response is elicited is the ‘set-point’. There are three types of arousal:
● Alarm arousal (external stimulation), for example, low temperature: a hibernating hedgehog maintains TB at 5°C. Lowered temperatures result in either increased metabolic rate or arousal.
● Periodic arousal (internal stimulation), for example lowered glucose levels.
● Final arousal: the rate is related to the species, varying from 30 minutes to many hours. It is noted that near arousal time, the temperature set-point fluctuates (for unclear reasons); the anterior of the animal warms before the posterior. In many species, both white adipose tissue and brown adipose tissue (BAT: see above, strategically situated between shoulder blades near the heart) aid warming.