Viral Pathogenesis
Acute cytolytic infection
The most obvious potential outcome of virus infection of a cell is death of the infected cell. This is referred to as a cytolytic or cytocidal infection. In order to maximize production of virus components, many viruses shut off host cell macromolecular synthesis. In order to keep their integrity, cells need to replenish damaged proteins, and cessation of this process will severely damage the cell. The accumulation within a cell of thousands of viral particles can have a considerable effect on the cell’s architecture. The ultimate outcome of these and other processes is the cell’s death. The explosive release of huge numbers of viral particles, which can then infect neighboring cells, results in lysis of the cell membranes.
Within several hours of the initial infection, this cycle may be completed. It is not difficult to find proof of cellular damage due to virus infection. Viruses can be grown in vitro in the culture of cells. As the virus replicates, light microscopy of these cell cultures reveals morphological modifications within the cells. Such modifications are referred to as a cytopathic (CPE) effect. Thus although viruses can not be visualized using a light microscope themselves, their effect on cells can be, and when using virus isolation in cell culture, diagnostic laboratories take advantage of this to prove that a patient is infected with a specific virus. Different viruses produce different CPEs. With experience, a highly educated guess can be made as to which virus is present within a specific culture that shows a CPE.
Virus-induced CPE is also present in vivo. Histological sections of virus-infected tissues may reveal the presence of multinucleate giant cells or the inclusion of bodies within cells consisting of rafts of viral particles). An interesting dilemma is created by the reliance of the virus on the host cell for replication. Although the virus is ultimately released from the cell once the cell is killed, before adequate replication of the virus has taken place, it is not in the interest of the virus to kill the cell too quickly. There is an additional complication of the host immune response within a normal host, one of whose primary roles is to destroy virally infected cells as quickly as possible to minimise the production of new viral particles. The patient’s end result will depend on the amount of infection-induced cell death and also on the degree of inflammatory response to the infection.
This will depend on a number of factors, including the dose of virus infection, the speed of virus replication and virus-induced cell death, and the speed and quality of host immune response. In many virus infections, the amount of cell death within the target organ is insufficient to impair the function of the organ or cause any symptoms to the patient. In most immunocompetent host infections, the immune system succeeds in eliminating the replicating virus and the patient survives. Sometimes the amount of virus-induced or immune-mediated cell death is so high that organ function is impaired and the patient is affected by acute symptomatic infection, e.g. acute hepatitis. At its extreme, this can lead to acute organ failure, e.g. fulminant hepatic failure may result from over-infection with the hepatitis B virus.
Chronic infection
Another form of virus-cell interaction is chronic or persistent infection, in which the virus is constantly produced and released from the infected cell, but the cell is capable of survival. Virus production continues for the natural life of the host cell. A patient with a chronic virus infection may have no symptoms at all due to their infection—such an individual is known as an asymptomatic (or healthy) carrier of the virus. However the disease may arise from chronic infection of the virus in one of two ways.
- Viral antigens are expressed on the cell surface as viruses replicate within cells. The host immune system in particular the T lymphocytes, is able to recognise these antigens as foreign and to provide an inflammatory response against the infected cells. If for whatever reason, the response fails to eliminate the virus-infected cells, the patient will suffer from a chronic inflammatory condition within the organ concerned. Thus in chronic hepatitis B infection of the liver, a large proportion of liver damage is actually caused by the host immune response to infected hepatocytes, rather than by any virus-induced cell death.
- All cells have to perform certain functions simply in order to remain alive. Collectively, these are called housekeeping functions. In addition, many cells have specialised functions, also known as luxurious functions. In chronic infection, by definition, the virus must not interfere with the cell’s housekeeping functions (otherwise the cell would die).However the presence of the virus may affect the cell’s luxurious functions, which may lead to clinical disease. When the newborn mice are infected with the lymphochoriomeningitis virus (LCMV) and followed over the next several months, the mice will not grow. LCMV infects the pituitary gland in these animals but does not cause cell death or inflammatory reactions. However the virus is present in growth hormone-producing cells and interferes with the production and secretion of the hormone—hence the failure of the mice to develop. To date, there is no human disease whose pathogenesis fits this model.
Latent infection
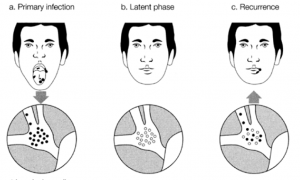
In the case of a latent infection, the viral genome is present within the cell, but there is no replication of the genome and few, if any, viral proteins are synthesized. The presence of a latent virus within the cell causes no harm to the cell. However a latent virus may be reactivated in response to certain stimuli. The virus then replicates, and this may have consequences for the cell and the body as a whole. All herpesviruses are subject to latency. After a primary infection, the virus will remain within that individual for the rest of their lives. Mostly the virus is in a latent state, but reactivation and subsequent disease may occur. Such reactivated or secondary infections are usually relatively trivial and highly localised, as unlike the very first or primary infection, which may be quite severe, the virus is reactivating in a host whose immune system has seen it before.
The herpes simplex virus (HSV) may therefore infect the oral cavity to cause orolabial herpes or gingivostomatitis (Fig. 1), which may or may not be symptomatic. Virus enters the nerve terminals (from the trigeminal nerve) and travels up the axons to reach the cell body (or ganglion) which is the site of HSV latency. Subsequent reactivation of the trigeminal ganglion virus causes the virus to travel back to the periphery where it causes one or a small cluster of vesicles known as cold sores.
Transformation
Transformation is a process in which cell infection leads to uncontrolled cell division, resulting in an immortal cell line. For example, in vitro Epstein-Barr virus infection of human B lymphocytes results in stimulation of cell division and the establishment of a continuous lymphoblastoid cell line. The fact that viruses can transform cells raised a clear question as to whether viruses can cause cancer. The answer is a clear yes.’ A number of virus infections have been shown to be closely related to the pathogenesis of a variety of malignant diseases.

YOU MAY LIKE:
REFERENCES: