Bacterial Cell Transport
- Every living cell requires from its surroundings raw materials for biosynthesis and energy production and must release to its environment the by-products of metabolism.
- A few non-polar compounds can dissolve in the lipid bilayer and cross the membrane unassisted, but for polar or charged compounds or ions, a membrane protein is essential for trans-membrane movement.
- In some cases, a membrane protein simply facilitates the diffusion of a solute down its concentration gradient, but transport often occurs against a gradient of concentration, electrical charge, or both, in which case solutes must be “pumped” in a process that requires energy.
- Ions may also move across membranes via ion channels formed by proteins, or they may be carried across by ionophores, small molecules that mask the charge of the ions and allow them to diffuse through the lipid bilayer.
- With very few exceptions, the traffic of small molecules across the plasma membrane is mediated by proteins such as trans-membrane channels, carriers, or pumps.
a) Passive diffusion
The movement of molecules across a membrane by passive transport does not require an input of metabolic energy. The molecule moves from a high concentration to a lower concentration. Passive transport by simple diffusion does not require the presence of integral membrane proteins. The rate of movement of a molecule (e.g. water, gases, and urea) by simple diffusion is directly proportional to its concentration gradient across the membrane.
B) Facilitated diffusion
- Passive transport by facilitated diffusion requires the presence of specific integral membrane proteins to facilitate the movement of the molecule (e.g. glucose, other sugars, amino acids, etc) across the membrane.
- The transport protein (e.g. the erythrocyte glucose transporter) is specific for a particular molecule, is saturable, displays binding kinetics, and is influenced by the temperature, pH, and inhibitors.
- The facilitated (or carrier-mediated) diffusion of a molecule across a biological membrane is dependent on specific integral membrane proteins, often called uniporters.
- The molecule binds to the protein on one side of the membrane; the protein then undergoes a conformational change, transports the molecule across the membrane, and then releases it on the other side.
- Molecules transported across membranes include hydrophilic molecules such as glucose, other sugars, and amino acids.
- The transport proteins are specific for one particular molecule or a group of structurally similar molecules. The transport proteins are capable of being saturated and are influenced by temperature, pH, and inhibitor molecules.
C) Active Transport
Active transport occurs against the concentration gradients and is mediated by carrier proteins. Metabolic energy is used to move ions or molecules against the concentration gradient. Active transport results in the accumulation of solute on one side of the membrane. Active transport is of two types’ Primary active transport and secondary active transport.
Secondary active transport
Secondary active transport is also known as indirect transport. It is the carrier protein-mediated transformation of molecules and ions. It occurs when the endergonic (uphill) transport of one solute is coupled with the exergonic (downhill) flow of different solute that was originally pumped by primary active transport. Secondary active transport is either symport or antiport, depending on whether two solutes move in the same or opposite direction.
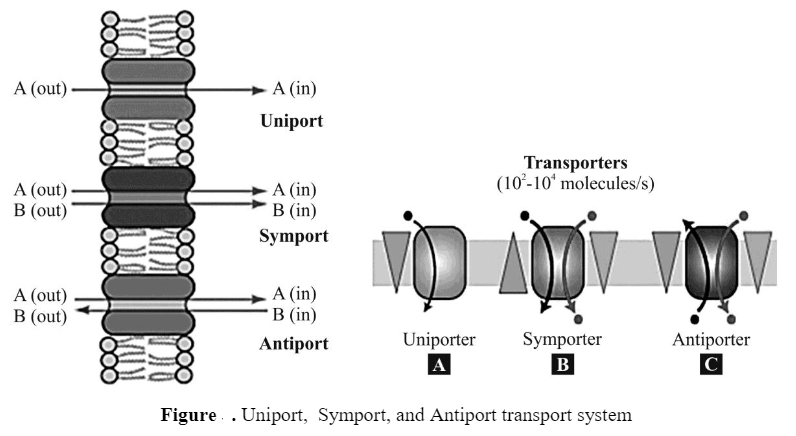
Concept of uniport, symport, and antiport
These are types of active transport that need carrier protein for transformation molecules from one side to another side of the membrane.
Symport – When the transported molecule and co-transported ion move in the same direction, the process is called symport.
Antiport – When the transported molecule and co-transported ion move in the opposite direction, the process is called antiport.
Uniport – Transporters carry only one substance from one side to the other side of the membrane that is known as uniport.
 |
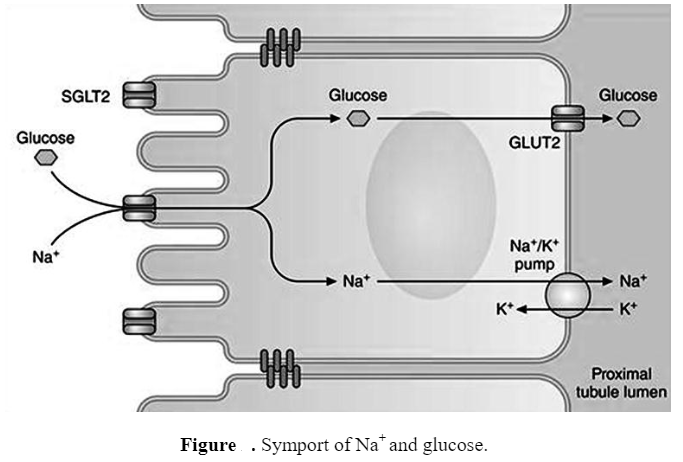
| Na+-glucose transporter: An example of secondary active transport |
- A common example of secondary active transport is the symport of Na’ and glucose. The trans-membrane protein Na-glucose transporter also known as Na -glucose co-transporters or symporters allows Na and glucose to enter the cell together.
- Na -glucose symporters are a family of glucose transporters present on the apical surface (facing the lumen) of the epithelium cell of the small intestine and actively transports glucose molecules into the cell from the gut.
- The Na+ flows down their concentration gradient while the glucose molecules are transported against their concentration gradient into the cell.
- Later the Na+ is pumped back out of the cell by the Na+-K+ ATPase and thus maintaining the inward Na+ gradient. The GLUT confined to the basolateral (basal and lateral) surfaces of the cell allows the same molecule to leave the cell by facilitated diffusion into the extracellular fluid on the other side of the epithelium.
- Similarly, lactose permease (also known as galactoside permease) present in the plasma membrane of the bacteria such as E. coli, utilizes the proton gradient across the membrane to co-transport proton and lactose (lactose proton symport).
D) Group translocation
- Inactive transport, solute molecules move across a membrane without modification. Many prokaryotes also take up molecules by group translocation, a process in which a molecule is transported into the cell while being chemically altered.
- For example, Phosphoenolpyruvate: Sugar phosphotransferase system (PTS). It transports a variety of sugars while phosphorylating them using phosphoenolpyruvate (PEP) as the phosphate donor.
PEP + Sugar (outside) —> Pyruvate + Sugar-P (inside)
- In E. coli and Salmonella typhimurium, it consists of two enzymes and a low molecular weight heat-stable protein (HPr). HPr and enzyme I (EI) are cytoplasmic.
- Enzyme II (EII) is more variable in structure and often composed of three subunits or domains. EIIA is cytoplasmic and soluble. EIIB also is hydrophilic but frequently attached to EIIC, a hydrophobic protein that is embedded in the membrane.
- A high-energy phosphate is transferred from PEP to enzyme II (EII) with the aid of enzyme I (EI) and HPr. Then a sugar molecule is phosphorylated as it is carried across the membrane by enzyme II (EII). Enzyme II (EII) transports only specific sugars and varies with PTS, whereas enzyme I (EI) and HPr are common to all PTS’s.
- PTS’s are widely distributed in prokaryotes. Aerobic bacteria lack PTS’s. Genera Escherichia, Salmonella, Staphylococcus, and other facultative anaerobic bacteria have phosphotransferase systems: some obligate anaerobic bacteria (Clostridium) also have PTS’s. Many carbohydrates are transported by these systems. Z.colitakes up glucose, fructose, mannitol, sucrose, N-acetylglucosamine, cellobiose, and other carbohydrates by group translocation.