History of Chromosome
- German biologist Walter Flemming in the early 1880s revealed that during cell division the nuclear material organizes itself into a visible thread-like structure which was named as chromosomes and stains deep with basic dyes.
- The term chromosome was coined by W. Waldeyer in 1888. Chrome is colored and soma is the body, hence they mean “colored bodies” and can be defined as higher-order organized arrangement of DNA and proteins.
- Chromosomes are structures that contain genetic material. It contains many genes or hereditary units, regulatory elements, and other nucleotide sequences. Chromosomes also contain DNA-bound proteins, which serve in packaging the DNA and control its functions.
- Chromosomes vary both in number and structure among organisms. „ They are complexes of DNA and proteins. The genome comprises all the genetic material that an organism possesses.
- In bacteria, it is typically a single circular chromosome while in eukaryotes, it refers to one complete set of nuclear chromosomes. The number of chromosomes is characteristic of every species.
- Benden and Bovery in 1887 reported that the number of chromosomes in each species is constant. W.S. Sutton and T. Boveri in 1902 suggested that chromosomes are the physical structures that acted as messengers of heredity.
Size and shape of Chromosome
- The size of the chromosome is normally measured at mitotic metaphase and may be as short as 0.25μm in fungi and birds to as long as 30 μm in some plants.
- However, most mitotic chromosome falls in the range of 5μm in man and 8- 12μm in maize. The monocots contain large-sized chromosomes as compared to dicots. Organisms with less number of chromosomes contain comparatively large-sized chromosomes.
- The chromosomes in the set vary in size. The shape of the chromosome changes from phase to phase in the continuous process of cell growth and cell division.
- During the resting/interphase stage of the cell, the chromosomes occur in the form of thin, coiled, elastic, and contractile, thread-like stainable structures, the chromatin threads.
- In the metaphase and the anaphase, the chromosome becomes thick and filamentous. Each chromosome contains a clear zone, known as centromere or kinetochore, along their length.
- Chromosomes may differ considerably in size and maybe categorized into groups. For example, in humans, the chromosomes are placed into seven groups from A to G. The largest chromosome is placed in group A and the smallest chromosome in group G. Between organisms, the size differences of chromosomes can be many fold and such differences even exist between species.
- Chromosomes can be observed inside the nucleus as thread-like structures that become visible under the light microscope when the cell is stained with certain dyes.
- The number of chromosomes in each cell of a particular species is always constant. Chromosomes are found to exhibit a characteristic splitting behavior in which each daughter cell formed by cell division receives an identical complement of chromosomes.
- There is a close relationship between the chromosomes and the DNA as the chromosomes contain DNA and proteins. While the amount of DNA per cell is constant, the amount and kinds of chromosomal proteins differ greatly from one cell type to another.
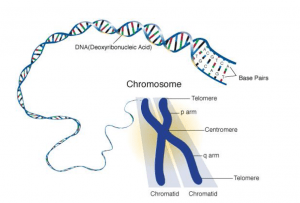
Structure of Chromosome
- A chromosome at mitotic metaphase consists of two symmetrical structures called chromatids.
- Each chromatid contains a single DNA molecule and both chromatids are attached to each other by the centromere and become separated at the beginning of anaphase.
- The spindle fibers act as molecular strings and are attached to chromosomes during cell division at a specialized region called the centromere. It appears as a constriction that divides the chromosome into two arms ( the shorter is called p and the larger is called q ).
- Centromere in a chromosome contains specific DNA sequences with special proteins bound to them, forming a disc-shaped structure, called the kinetochore.
- In electron microscope, the kinetochore appears as a plate or cup-like disc, 0.20-0.25 nm, in diameter situated upon the primary constriction or centromere.
- The chromosomes of most organisms contain only one centromere and are known as monocentric chromosomes.
- Telomere is the chromosomal end that prevents other chromosomal segments to be fused with it. Besides the primary constrictions or centromeres, chromosomes also possess secondary constriction at any point of the chromosome and are constant in their position and extent.
- These constrictions are helpful in identifying particular chromosomes in a set. Chromosomes also contain nucleolar organizers which are certain secondary constrictions that contain the genes coding for 5.8S, 18S, and 28S ribosomal RNA and induce the formation of nucleoli.
- Sometimes the chromosomes bear round, elongated, or knob-like appendages known as satellites. The satellite remains connected with the rest of the chromosomes by a thin chromatin filament.
Chromosome number
- There are normally two copies of each chromosome present in every somatic cell. The number of unique chromosomes (N) in such a cell is known as its haploid number, and the total number of chromosomes (2N) is its diploid number.
- The suffix ‘ploid’ refers to chromosome ‘sets’. The haploid set of the chromosome is also known as the genome. Structurally, eukaryotes possess large linear chromosomes, unlike prokaryotes which have circular chromosomes.
- In Eukaryotes other than the nucleus chromosomes are present in mitochondria and chloroplast too. The number of chromosomes in each somatic cell is the same for all members of a given species.
Autosome and Sex Chromosome
In a diploid cell, there are two of each kind of chromosome (termed homologous chromosomes) except the sex chromosomes. In humans, one sex has two of the same kind of sex chromosomes and the other has one of each kind. In humans, there are 23 pairs of homologous chromosomes (2n=46). The human female has 44 nonsex chromosomes, termed autosomes, and one pair of homomorphic sex chromosomes given the designation XX. The human male has 44 autosomes and one pair of heteromorphic sex chromosomes, one X and one Y chromosome.
Centromere
- Centromere refers to the most prominent region of condensed mitotic chromosomes called the primary constriction. This region was initially called centromere as it was invariably located in the middle between the ends of two chromosome arms. Later, the term was extended to describe the primary constriction of all mitotic chromosomes even when it is not located in a central position.
- Centromeres have multiple roles during mitosis and are responsible for the accurate segregation of the replicated chromosome during mitosis and enabling the equal distribution of genetic material during cell division.
- When chromosomes are stained they typically show a dark-stained region that is the centromere. The actual location where the attachments of spindle fibers occur is called the kinetochore and is composed of both DNA and protein.
- On either side of the centromere, is a trilaminar plate structure called the kinetochore. It is a multiprotein complex located at the surface of the chromosomes that binds spindle microtubules and regulates chromosome movement in mitosis.
- It is also the final site of sister chromatid pairing before segregation takes place. The centromere and the kinetochore help in ensuring the proper orientation of chromosomes at metaphase.
- Microtubules are attached to the kinetochores. Though centromeres have been conserved throughout evolution, they show structural variability and are classified into two different types.
- Centromeres may be diffused as is found in many arthropods and plants (Luzula) or localized as are generally present in eukaryotes. In diffused centromeres, spindle microtubules attach along the entire length of the chromatids while in localized centromeres, there is a single region of attachment for spindle microtubules.
Telomeres
- The ends of linear chromosomes are called telomeres. These specialized structures play important role in protecting the ends of chromosomes from attack by nuclease enzymes.
- They also prevent the chromosomes from joining together as the broken chromosomes attach immediately. Moreover, the extreme 5′- terminus of a linear DNA molecule comprises an RNA primer that is not replaced with DNA.
- A telomere is specialized to make the natural end of a linear chromosome behave differently from a simple double-stranded DNA break.
- Telomeres, or ends of eukaryotic chromosomes, contain no genes. Instead, they are composed of many repeats of short, GC-rich sequences. The exact sequence of the repeat in a telomere is species-specific.
- These repeats are added to the very 3′-ends of DNA strands, by semi-conservative replication by telomerase. Telomeric DNA is made up of multiple copies of a short sequence, 5′-AGGGTT-3′ in humans which is repeated possibly a thousand times or more at the extreme ends of each chromosomal DNA molecule.
- The actual structure is different for each telomere. 5′-3′ strand is G rich while 3′-5′ strand is C rich. The repeat sequences act as binding sites for telomeric-specific proteins.
- The bound proteins probably act as a cap, preventing the ends of the chromosomes from being degraded or fusing with other chromosomes.
- The telomerase enzyme consists of a protein subunit and an RNA molecule. Telomerase RNA contains at one position a short sequence identical to one or more repeat sequences of the Crich strand of the telomere.
- It acts as a template for the synthesis of a repeat sequence of G-rich strands. Thus the shortening effect of DNA replication can be counterbalanced by repeatedly extending the G-rich strand.
- The telomere is a very important part of a chromosome. It is required for the complete replication of the chromosome. It forms caps that protect the chromosome from nuclease. It prevents the ends of chromosomes from fusing with one another. It facilitates interaction between the ends of the chromosome and the nuclear envelope in some types of cells.
References: