Definition
According to “Balinsky”, the process of gastrulation is one of displacement of part of blastoderm of an early embryo. As a result, the endodermal and mesodermal organ rudiment are removed from the surface of the blastula and brought into interior of the embryo, where the respective organs are found in the differentiated animal. Thus, the single layer of cells, the blastoderms given rise to three germ layers 1.e. ectoderm, endoderm and mesoderm.
Gastrulation may be defined as the dynamic process during with major presumptive organ forming areas of blastula becomes arranged and recognized in way which permits their ready conversion of body plan of particular species.
Major events of gastrulation:
1) The rhythm of cellular division is slowed down.
2) A rearrangement of blastomeres of embryo by means of morphogenetic movements.
3) The type of metabolism changes and the rate of oxidation is intensified.
4) The nuclei become more active in controlling the activities of embryonic cells.
5) During gastrulation chemo differentiation is started by the synthesis of new and different kinds of protein molecules.
Basic cell movement in gastrulation:
Gastrular Morphogenetic Movements
Gastrulation is a nicely integrated, dynamic process which is controlled largely by intrinsic forces bound up in the specific physico-chemical conditions of the various presumptive organ forming areas of the late blastula and early gastrula. These internal forces in turn are correlated with external conditions. In most vertebrates. it includes different kinds of morphogenetic movements.
The movement of the blastomeres from one places of the embryo the another to establish a particular form or structure is a common embryological procedure. This type of cell movement is described as formative or morphogenetic movement, because, it results in the generation of a particular form or structural arrangement (Vogt, 1925, Trinkaus, 1969 and Berrill. 1971).
Such morphogenetic movements occur in the embryo during blastulation, gastrulation, tubulation and organogenesis. They do not occur only during the embryonic stages, but also in an adult animal. But, there is a fundamental difference in the two, that the morphogenetic movements of an adult animal are of a reversible nature, while, the morphogenetic movements of gastrula, etc., are of irreversible nature, i.e., each part remains in the position into which it has been brought by the preceding movements.
The following two basic types of morphogenetic movement patterns are involved in gastrulation:
1. Epiboly; 2. Emboly.
1. Epiboly:
The word epiboly is derived from the Greek, meaning a “throwing on” or “extending upon”. It represents the natural tendency of epithelial sheet to expand into free areas over a substratum. Epibolic sheets are usually outside, or ectodermal, and tend to envelope and surround inner sheets. Sometimes sheets expand toward a single point on the surface. This type of epibolic movement is called convergence (see Grant, 1978).
Epiauxesis:
In Ctenophora, in various spirally cleaving eggs (e.g., Annelida and Mollusca), and in some vertebrata (Pisces), a modified type of epibolic movement called epiauxesis occurs (Lovtrup, 1974). Four conditions must be fulfilled for epiauxesis to occur, namely, absence of a hyaline membrane, presence of a surface to which the blastomeres adhere and on which they may spread, a certain intercellular adhesion and that they are mitotically active. In Annelida and Mollusca epiauxesis involves overgrowth of ectodermal (micromeres) over yolk-laden vegetal pole cells (macromeres). Overgrowth of micromeres not only depends upon the subdivision of the existing micromeres, but also upon the addition of new ones budding off from macromeres.
Convergence:
Convergence is the actual migration of cells from the outer surface of the blastula to the external margin of the blastoporal lip. During gastrulation the process of convergence and involution, go simultaneously. If the involution is not there, the converging cell would tend to pile up along the outer edge of blastopore lips.
In frog most of the chorda mesodermal blastomeres 1st converge to the dorsal blastoporal lip and then rotate inwardly to occupy the position just below the ectodermal blastomeres.
2. Emboly:
This is just opposite to epiboly. In emboly, sheets fold in or invaginate; that is, a sheet migrates inside a cell aggregate or beneath another layer because of local changes in cell shape or motility. Embolic movements include invagination, involution and ingression, movements displacing future or prospective endoderm and mesoderm into the interior of the embryo.
1) Invagination: It is the sinking in (in pocketing) of a layer of cells and is similar to punching in of an expanding layer at a specific point. The reverse of invagination is evagination.
2) Involution: It is turning in of an expanding layer at a specific point. In involution, a surface layer of cells turns inside itself and continues to expand along the inner surface of the superficial layer. Thus, in involution, cells which are at some distance from the point of invagination, move to the margin of the site of invagination (e.g., at a lip of blastopore during amphibian gastrulation), flow over it and move inside.
3) Ingression or poly invagination: In this type of emboly, small groups of independent cells separate from a primary epithelial layer and migrate into the blastocoele cavity or other embryonic spaces as a new layer. For example, in primitive streak of amniote embryos, cells may sink inside as individuals, changing their relations to each other in the process.
Infiltration: It occurs during the development of endoderm in amniotes. Individually cells detached themselves from the surface of blastoderm and fall into primary blastocoels. Here they arrange themselves to form a layer of endoderm cells.
Delamination is a type of ingression that forms an inside sheet by mass separation of a layer of cells from an epithelium. Sometimes, this process is accomplished by division of cells in a plane parallel to a tissue space.
Divergence:
It is opposite of convergence. For example, after cells have involuted over the blastoporal lips during gastrulation, they migrate and diverge to their future positions within the developing embryo. Such divergence occurs in lateral plate and ventral mesoderm in frog; and in lateral plate and extra-embryonic mesoderm in reptiles, birds and mammals.
Gastrulation in Egg:
Gastrulation in an Isolecithal Egg
- The main processes involved in the conversion of a monoblastic blastula into a diploblastic and stratified gastrula are invagination, involution, and a kind of epibolic morphogenetic movements of different blastomeres with the flattening of blastoderm at the vegetal pole and subsequent inward bending.
- The embryo, instead of being spherical, becomes converted into a cup-shaped structure with a large cavity in open communication with the exterior on the side that was originally the vegetal pole of the embryo.
- The cup has a double wall-the external and internal epithelial layers which are continuous with each other over the rim of the cup-shaped embryo.
- The external lining consists of ectoderm i.e., the presumptive epidermis and presumptive nervous system. The internal lining consists mainly of the endoderm. Very soon, the presumptive material for notochord and mesodermal crescent slide-in or involute so as to occupy a position on the internal wall of the cup.
- In this way, the endoderm, the mesoderm, and the notochord material disappear from the surface of the embryo into the interior where they belong. The external surface of the embryo now consists of an ectoderm.
- The embryo in this stage of development is called a gastrula. The cavity of it (formed due to invagination of endoderm and mesoderm) is called primary gut or archenteron and its opening to the exterior is called the blastopore.
- The blastopore denotes the pathway by which the endoderm and mesoderm pass into the interior of the embryo. Its rims, therefore, are usually referred to as the lips of the blastopore the dorsal Lip, the ventral lip, and the lateral lips.
- At the initial stage of gastrulation, the blastopore 1s very broad but soon the lips of the blastopore begin to contract so that the opening which leads into the archenteron becomes smaller and is reduced to an insignificant fraction of the original orifice.
- This contraction of the lips of the blastopore is connected with the marked increase in the mitotic activity which occurs in the cells of the presumptive epidermal and neural ectoderm, notochord, and mesoderm and they’re spreading out by epibolic morphogenetic movements as well as the disappearance of the mesodermal crescent material and the presumptive notochord from the rim of the cup-shaped embryo.
- As more material is shifted into the interior of the gastrula, the remnants of the blastocoel become completely obliterated by the two walls of the embryo coming in contact with each other.
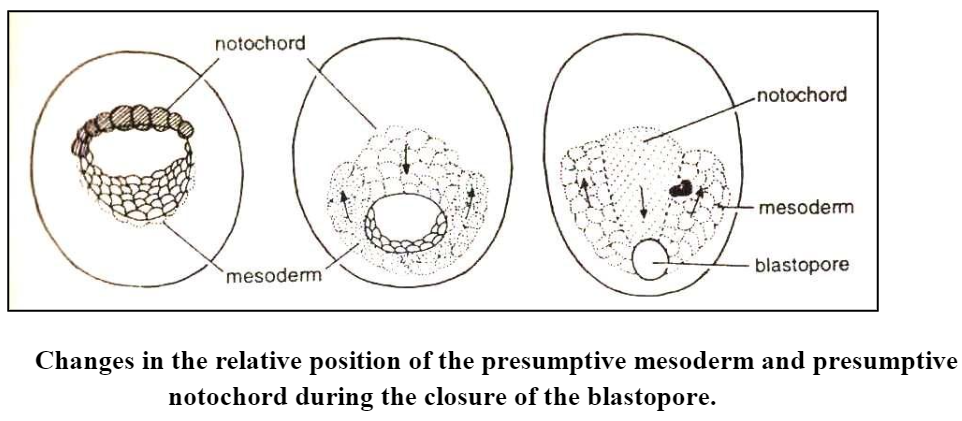
- As the presumptive notochord and the mesodermal crescent shift into, the interior of the gastrula, they also change their position relative to each other. In the blastula, these two areas lie on opposite sides of the embryo.
- Now the lateral horns of the mesodermal crescent converge toward the dorsal side of the embryo and come to lie on both sides of the presumptive notochord. Now due to contraction of the rim of the blastopore, the embryo becomes elongated in the anteroposterior direction, and all the various presumptive areas participate in this elongation.
- The elongation of the notochord and mesodermal material brings them into still closer contact with each other, the notochordal material shifting backward, in between the two horns of the mesodermal crescent material.
- As a result of these movements, the notochordal material becomes stretched into a longitudinal band of cells lying medially in the dorsal inner wall of the gastrula and flanked on both sides by bands of mesodermal cells similarly stretched in a longitudinal direction. The remainder of the lateral, ventral, and anterior parts of the inner wall of the gastrula consists of endodermal cells.
- The external wall of the gastrula similarly takes part in the elongation of the embryo. One of the results of this is that the presumptive material of the nervous system becomes stretched into a longitudinal band of cells lying mediodorsally over the notochordal material but being somewhat broader than the latter.
Gastrulation in a Moderately Telolecithal Egg
- In a moderately telolecithal or mesolecithal egg, the cellular movements during gastrulation can be best described as follows :
- A mid-sagittal section through the blastula of the moderately telolecithal egg is shown in (Fig. A). Numbers | to 9 represent specific regions of the blastoderm of the animal hemisphere, with number 5 occurring at the animal pole.
- Now, because the cells in the region of 5 are reproducing at such a fast rate, relative to the vegetal cells simply because the blastomeres in the vegetal hemisphere still contain large quantities of mitosis-inhibiting yolk.
- Now the fastly reproducing cells must either accumulate or “flow down” over the ventral surface of the blastula (Fig. B).
- This flowing of cells is referred to as epiboly and is very instrumental in the formation of gastrula. At one point of the egg’s surface the cells cease their downward movement and begin to flow into the blastula itself. When a layer of cells grows into a cavity rather than simply pushing into it, the process is called involution.
- This combination of epiboly and subsequent involution creates a number of new morphological structures. The new cavity which is beginning to form is the gastrocoel or archenteron.
- The opening into this cavity is the blastopore. As development of thegastrocoel proceeds, more and more of the cell indicated by | to 4 will involute, until eventually the configuration shown in Fig. E is reached.
- The blastocoel has been nearly obliterated by the over-growing archenteron. The cells represented by 6 to 9, however, are not stopped in their progression toward the vegetal pole but continue around the egg until they also approach the blastopore from the ventral side. Here they also involute, forming the ventral lip of the blastopore.
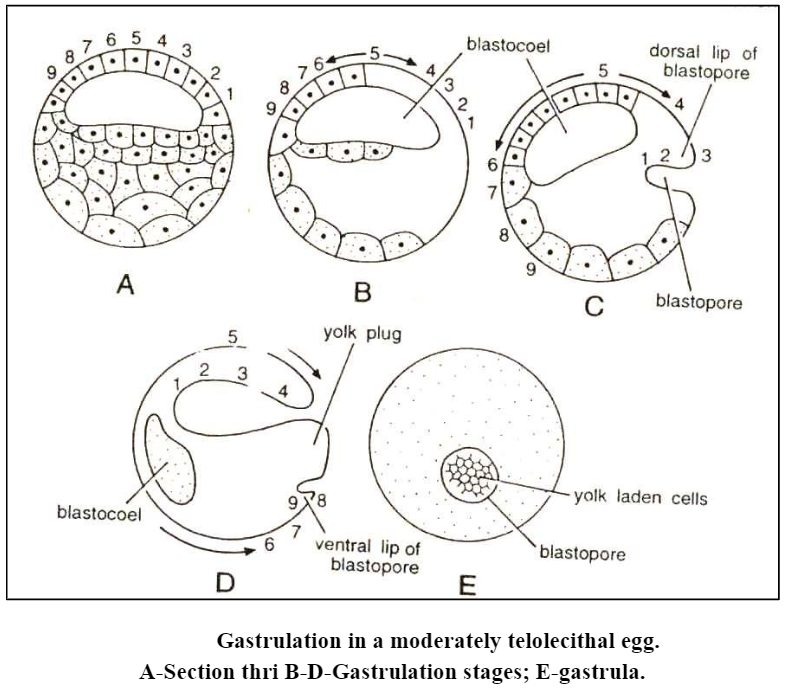
- As a result of the cellular movements, the entire portion is occupied by the blastopore. Through the blastopore, the yolk-laden blastomeres of the original ventral surface of the blastoderm can be seen. It is for this that the stage is called the yolk-plug stage.
- In general, amphibian gastrulation may be summarized as follows:
- The superficial endodermal cells of the marginal zone sink into the deep endoderm as bottle cells and the archenteron begins to form.
- Prospective mesoderm spreads on the highly adhesive lower surface of the overlying ectoderm.
- The spreading ectoderm cells at the same line expand as a sheet over the invaginating mesoderm and replace it as it disappears from the surface.
Gastrulation in a Heavily Telolecithal Egg
- The inert yolk mass is proportionally so enormous that it cannot participate even passively, in the gastrulation process. For this reason, the events of gastrulation are confined to the blastoderm alone which contains cells of all the three future germ layers.
- The gastrular morphogenetic movements which take place during gastrulation are the same as described for amphibians 1.e. convergence, involution, and elongation.
- In chicks, the two aspects of gastrulation, namely, the advance of the blastodermal margin and the establishment of a visible embryonic axis, are regionally two separate processes.
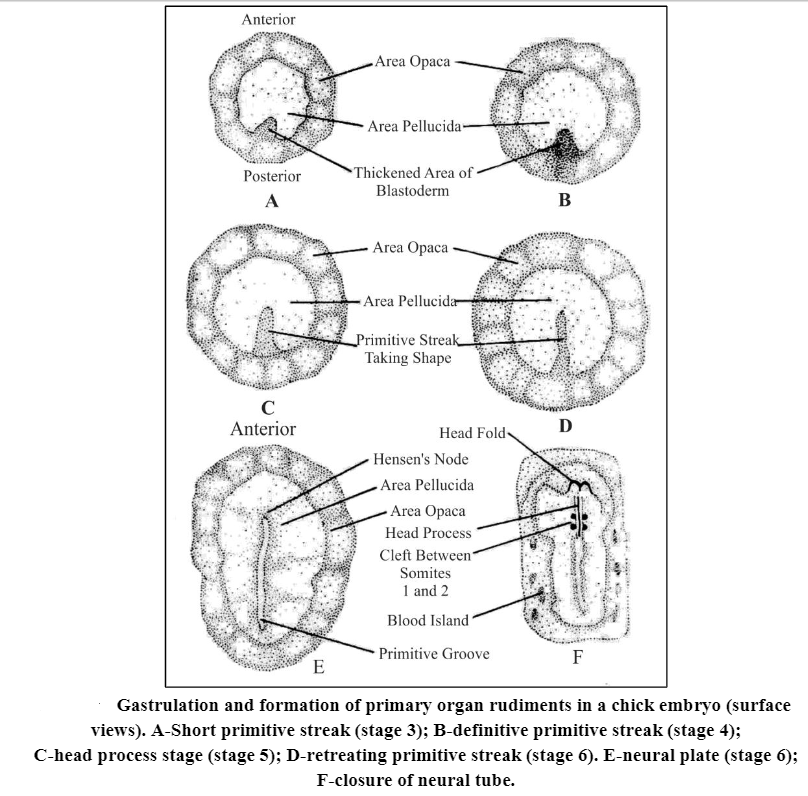
- The growth of the blastoderm margin is exclusively concerned with the enclosure of yolk within a yolk sac, while a primitive streak, first appears in the midline at the posterior quadrant of the area pellucida and then elongates anteriorly and becomes constricted.
- The elongated axis of the primitive streak foreshadows the anteroposterior axis of the future embryo.
- It has been suggested that the streak elongation is due to an incorporation of new material, added to one end or the other.
- The morphogenetic movements leading to the formation of the primitive streak begin as a movement of cells toward the posterior-median end of the blastoderm.
- Much of the posterior half of the area pellucida at the pre-primitive streak stage enters the primitive streak and becomes mesoderm.
- Some forward movement takes place in the middle line as the primitive streak forms. As soon as the streak appears, it is characterized by a typical bipolarity, i.e. the anterior half yields only extraembryonic mesoderm.
- As the definitive streak is formed a groove appears along its length, the primitive groove with the raised edges called the primitive ridges.
- It is toward this groove that cells migrate and via this groove that the cells reach the interior. The thickening of the blastoderm that constitutes primitive streak corresponds to the dorsolateral margins of the amphibian blastopore, here fused as a single structure.
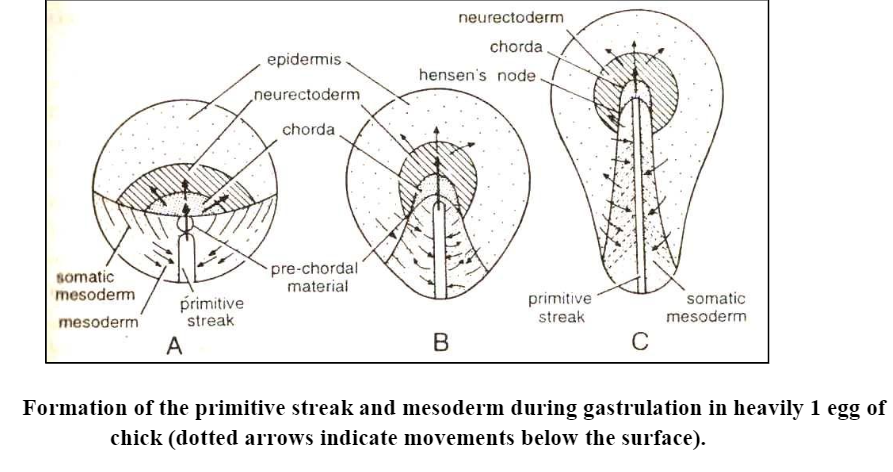
- The thickened anterior end of the streak is known as the primitive knot or Hensen’s node. corresponds to the dorsal lip.
- The primitive ridges correspond to the lateral lips. As in the amphibian, the lips are continually changing structures, being composed of cells in the process of turning in from the surface layer (epiblast) and moving into the interior. The movement of cells toward the streak and down into it is termed immigration.
- In birds and reptiles, the mesoderm, as well as the endoderm, invaginate through the primitive streak. The lower cellular layer of the blastoderm, however, comes from more than one source.
- Large blastomeres become progressively detached from the upper layer at a very early stage, while embryonic endoderm later invaginates from the primitive streak to a lower layer position.
- According to Spratt and Haas, the primitive streak forms as a result of the convergence of epiblast cells to the midline of the blastoderm. Here they sink in and migrate out laterally between the epiblast and the hypoblast to form the mesoblast.
- Some endoderm, as well as the mesoderm, originates from the epiblast, invaginating at the primitive streak. Migration of both the presumptive mesodermal and endodermal cells laterally outward and away from the primitive streak is a continuing process.
- Either new cells must enter by immigration from the epiblast or they may result from a relatively high rate of cell division within the streak.
- The evidence is contradictory. However, mesoblast cells are seen to be connected to each other and to cells of the primitive streak by junctional contacts, especially along their trailing surfaces. They do not break away from the primitive streak as free cells and migrate as individuals but form a loosely connected network.
- In general, the tissue sinks in along the centerline of the streak. It then moves laterally and forward beneath the overlying ectoderm and subjacent endoderm to form the chordomesoderm, which is equivalent to the chordamesoderm sheet of the amphibian embryo.
- The presumptive notochordal tissue of the chick underlines the middle of the embryonic axis, with lateral mesoderm at each side. As this continues, with such tissue moving forward to the limits of the embryo-to-be, the primitive streak tissue undergoes corresponding attrition at its anterior end. In effect, the primitive streak becomes progressively shorter and seems to shift posteriorly.
- The shortening of the primitive streak is accomplished by the posterior regression of Hensen’s node, which consists primarily of chordamesoderm in the process of turning inside. Once inside the tissue of the prospective notochord moves to underlie the epiblast in the region that will become the neural plate. As the most anterior parts of the embryonic axis are formed, the node progresses down the streak to produce more posterior parts.
- It is observed that differential growth occurs in the blastoderm, which exhibits radial and polarized patterns of differential rates of cell multiplication in various regions of the cell population.
- The outcome of differential growth is a ring-shaped, lower-level population-density- gradient with maximal density on one side where the primitive streak is normally initiated.
- The important property of the blastoderm is the presence of group of rapidly dividing cells located eccentrically. The cells of this growth centertend to spread out rapidly as a lower cell layer (hypoblast) along the underside of the radially symmetrical and immobile upper layer (epiblast).
- The spreading lower cells form a coherent sheet that moves like a viscous fluid away from its source until it reaches the marginal zone of the blastoderm, then the cells at the edge move circumferentially to the right to the left.
- The pattern of movement of the lower sheet of cells is accordingly like a fountain, and it appears to be entirely a consequence of the geometry of the blastoderm. In normal development, the axis of symmetry of the lower layer movement pattern and the position of the head-tail axis of the future embryo coincide.
- If the main axis of the movement pattern is experimentally shifted, the head-tail axis is correspondingly shifted. If the lower layer movement is blocked, no embryo develops. However, the concept of a single, embryo initiating, growth center may be due to the coordination of cells throughout the entire area pellucida rather than to a high rate of mitosis and density of cell population.
- Both the area pellucida endoderm and also mesoderm arise from the primitive streak. Endoderm formation rapidly outstrips that of the mesoderm, so that an endodermal layer is complete when the mesoderm is still only partly invaginated.
- This premature endoderm determines the orientation of the primitive streak. If the endodermis is removed from a blastoderm at an early primitive streak stage and is then replaced at right angles to its original position, the streak comes to form but bends at its anterior end toward the original anterior end of the endoderm.
- Therefore, the endodermal layer at least plays a part in directing primitive streak formation and the cell movements of the upper layer.
Significance of Gastrulation
(1) It brings the presumptive structures from a position on the external surface of the blastula into their normal positions in the embryo.
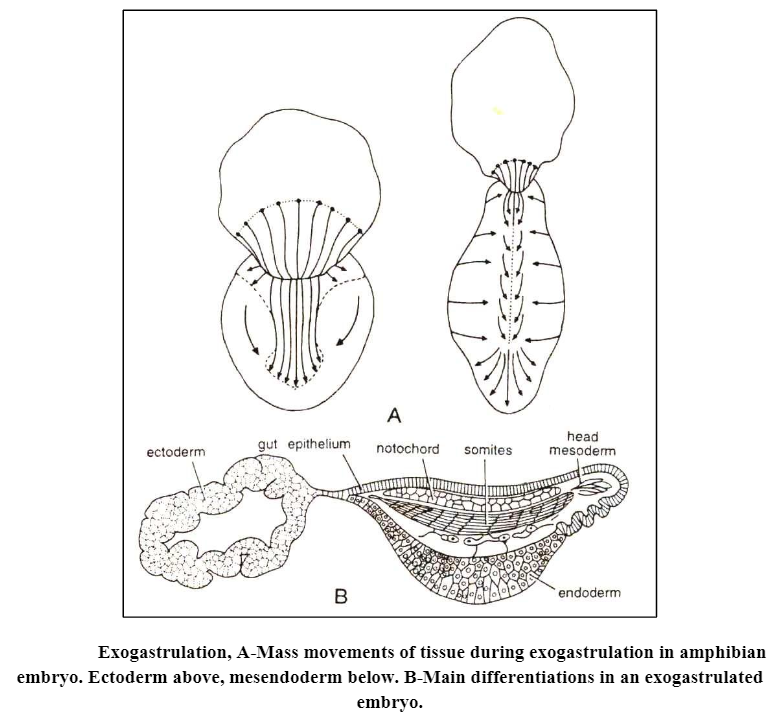
(2) It is necessary for the formation of the nervous system. This can be proved by producing abnormal gastrulae called exogastrulae of frogs. In some amphibian eggs, if the outer protective membranes are removed, the eggs form exogastrulae. In an exogastrula, the endodermal and mesodermal blastomeres evaginate (viz. grow outward) rather than invaginate and the mesoderm and endoderm are not covered with ectoderm, the ectoderm remains isolated from the rest of the tissues.
The important aspect of the exogastrulation is that endodermal and mesodermal structures develop to a great extent, but no nervous structures form. Neither the neural plate nor any of the sensory structures develop from the ectoderm if chorda mesodermal are a does not move inward and makes contact with the neural ectoderm.