How Genes Determine Traits?
We have seen that the central dogma is the key principle of molecular genetics:
- The sequence of nucleotides in the gene specifies the sequence of amino acids in the protein using the messenger RNA as an intermediary molecule in the encoding process.
- It is one of the irony of genetics (and a consequence of the biochemical complexity of organisms) that while the link between genes and proteins is conceptually simple, the link between genes and traits is certainly not simple.
- Most visible traits of organisms are the net result of many genes acting in combination with environmental factors. The relationship between genes and traits is therefore often complex for one or more of the following reasons:
1. More than one trait can be affected by one gene.
2. More than one gene can be affected by one trait.
3. Many traits are affected by environmental factors as well as genes.
Now let’s look at each of these principles, with examples.
Pleiotropy: One Gene Can Affect More Than One Trait
- A mutant gene may affect a number of apparently unrelated traits. The mutation is then said to show pleiotropy, and the various manifestations of the mutation are known as pleiotropic effects.
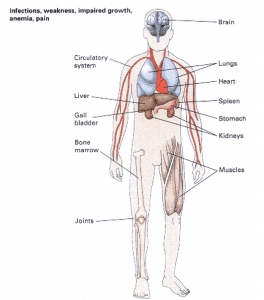
- An example of a mutation in human beings with multiple pleiotropic effects is sickle cell anaemia, which affects the major oxygen-carrying protein in the red blood cells. Major organs and organ systems affected by pleiotropic effects.
The mutation is shown in Figure 1 below:
- The underlying mutation in sickle cell anaemia is the β-globin gene, which codes for the β polypeptide chains present in the oxygen-carrying protein of the red blood cells. The molecular basis for the disease is shown in Figure 2 below.
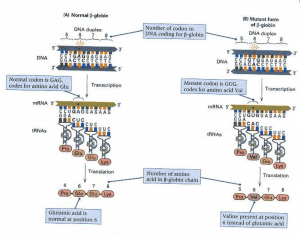
- Figure 2 shows the region of the β-globin gene that codes for amino acids 5 through 8 (the complete polypeptide chain is 146 amino acids in length). The sickle-cell mutation changes the base pair with the sunburst.
- As shown in Part B, the mutant form of the gene contains the T — base pair instead of the normal A — T base pair. As a result of the mutation, the mRNA contains the codon of the GUG. Because GUG Valine (Val) codes, this amino acid is incorporated into the polypeptide in place of the normal glutamic acid (Glu) at position 6.
- The defective β-polypeptide chain gives the haemoglobin protein a tendency to form long, needle-like polymers. Red blood cells in which polymerization occurs are deformed into a crescent, sickle-like shape.
- Some of the deformed red blood cells are destroyed immediately (reducing the oxygen-carrying capacity of the blood and causing anaemia), while others may clumsy and clogg blood circulation in the capillaries.
- The consequences of replacing Glu Val are a profound set of pleiotropic effects. All of these effects are related to the breakdown of red blood cells, the decreased oxygen-carrying capacity of the blood, or physiological adjustments that the body makes to try to compensate for the disease (such as spleen enlargement).
- Patients with sickle cell anaemia suffer from severe pain. Anemia is the cause of impaired growth, weakness, and jaundice. People affected are so generally weakened that they are susceptible to bacterial infections, which are the most common cause of death in children with the disease.Although sickle cell anaemia is a serious genetic disease that often leads to premature death, it is relatively common in areas of Africa and the Middle East where the type of malaria caused by the protozoan parasite Plasmodium falciparum is widespread.
- The association between sickle cell anaemia and malaria is not coincidental. It’s an infection. In the life cycle of the parasite, the bite of the mosquito passes from the mosquito to the human. The initial stages of infection take place in the liver cells, where specialised forms of the parasite are produced that are capable of infecting and multiplying red blood cells. Widespread infection of the red blood cells impairs the ability of the blood to carry oxygen, causing the weakness, anaemia and jaundice of malaria.
- However, in people with mutant β-hemoglobin, it is thought that the infected blood cells are sick and quickly removed from circulation. This controls the spread of the parasite among the red blood cells and reduces the severity of the malaria infection.
- There is therefore a genetic balancing act between the prevalence of sickle cell anaemia and the prevalence of parasitic malaria. If the mutant β-hemoglobin becomes too frequent, more lives will be lost from sickle cell anaemia than from malaria protection; on the other hand, if the mutant β-hemoglobin becomes too rare, fewer lives will be lost from sickle cell anaemia, but the gain will be offset by more malaria deaths.
Epistasis: One Trait Can Be Affected by More Than One Gene
- Each trait requires numerous genes for proper development, metabolism, and physiology. As a result, more than one gene can affect one trait. An example of this principle is shown in Figure 3 below, which shows the effects of two genes in eye pigmentation in Drosophila.
- Genes are: vermilion (v) and cinnabar (cn). These genes encode the enzymes V and Cn, respectively, that are used in the biochemical pathway that converts the amino acid tryptophan to the brown eye pigment xanthommatin through a series of intermediate substances I1, 12, and so on.
- Each step in the pathway is catalysed by a different enzyme encoded by another gene. The non-mutant, or wild, eye colour of Drosophila is a brick-like red because the pigment cells contain not only xanthommatin but also a bright red pigment called drosopterin, which is synthesised by a different biochemical pathway.
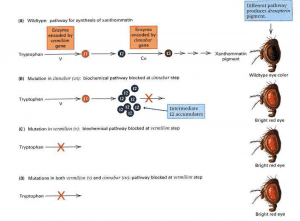
- As shown in Figure 3, flies that are mutant due to lack of xanthommatin. They’ve got bright red eyes because of the drosopterin. Flies mutant for cn have a non-functional Cn enzyme, so the pathway is blocked at the point where Cn should function.
- Since there is no functional Cn enzyme capable of converting the intermediate 12 to the next intermediate along the way, 12 accumulates in cn flies. Mutant v flies are also lacking xanthommatin, but for a different reason.
- The pathway is blocked in these flies because there is no functional V enzyme. It does not matter whether the enzyme Cn is present, because without the enzyme V, there is no 12 for Cn to work on. The pathway in flies with a mutation in both cn and v.
- The situation is the same as in flies with a v mutation only because the pathway is blocked at this stage due to the lack of a functional V enzyme. Epistasis is the general term for gene interaction. Freely translated from Greek, epistasis means “to stand over.”
- Epistasis means that the presence of one mutation “stands over” or conceals the effects of another mutation. In the example in Figure 3, we would say that v is epistatic to cn, because it is impossible to determine, from the status of the xanthommatin pathway, whether the cn gene is mutant or wild. The converse is not true: in flies with a mutation in the cn, the presence or absence of an intermediate 12 indicates whether the mutation in the cn is true.
- The V gene is mutant or wild. If 12 accumulates, the enzyme V (and the wild type gene v) must be present; whereas if 12 is missing, the enzyme V (and the mutant gene v) must be non-functional.
- An important feature of genetic terminology is also shown in the example in Figure 3. Although both vermilion and cinnabar are necessary for the synthesis of the brown pigment, the names of the genes are shades of bright red.
- It seems illogical at first, but mutant genes are named for their effects on the organism.Because mutations in either vermilion or cinnabar result in bright red eyes, the gene names make sense even though the products of both genes function in the brown-pigment pathway.
Effects of the Environment
- Genes and the environment are also interacting. In order to assess the interaction between genes and the environment, consider the “anaemia” trait, which refers to a generalised weakness due to an insufficient number of red blood cells or an insufficient amount of blood.
- There are a lot of different types of anaemia. Some forms of anaemia are genetically determined, such as sickle cell anaemia. Other forms of anaemia are caused by the environment; for example, anaemia caused by chronic dietary iron deficiency or malaria infection.
- Other forms of anaemia are caused by the combination of genetic and environmental factors. For example , people with a mutant form of the enzyme glucose-6-phosphate dehydrogenase (G6PD), an enzyme important for maintaining the integrity of the red blood cell membrane, become severely anaemic when they eat fava beans because the substance in the beans triggers the destruction of the red cells. The disease is called favism due to its association with fava beans.the disease is called favism, but a more common name is G6PD deficiency.
- Red cell destruction in people with G6PD deficiency may also be triggered by a variety of chemicals, such as naphthalene (used in mothballs) as well as certain antibiotics and other drugs.
- The G6PD deficiency, which mainly affects males, is relatively high in populations in coastal regions around the Mediterranean Sea. The defect in red blood cells is thought to increase resistance to malaria.
- With these examples as background, consider the following question: is anaemia caused by heredity or the environment? There is no simple answer to that. As we have seen, there are many possible causes for a complex trait such as anaemia. Some types are genetically determined, some are of environmental origin, and some require genes and the environment to be expressed.
- The gene-versus-environment issue is exceptionally clear in the case of anaemia only because the various forms of disorder have already been sorted out and the causes assigned, be they genetic or environmental, or both.
- However, before the different forms were distinguished, anaemia was considered to be a highly complex condition, and all varieties were lumped together. Without dividing the disorder into categories, all that could be concluded was that family history seemed to be important in some cases, but not all, and that the environment certainly played a role as well.
- Most complex traits are analogous to anaemia in that they consist of different conditions lumped together because of their overall similarity. A familiar example of this is heart disease. It is well known that hereditary risk factors for heart disease are related to the metabolism of saturated fats and cholesterol.
- Some rare diseases with a strong genetic component have already been identified. Environmental risk factors for heart disease include smoking cigarettes, overweight, lack of exercise, high dietary intake of saturated fats and cholesterol, and so on.
- In the population as a whole, the overall risk of heart disease is determined by both genetic and environmental factors, and some of the factors act synergistically, which means that the risk from two factors together is greater than would be predicted from the risk of each factor considered on its own.
- The example of heart disease also shows that genetic and environmental effects can be compensated for. For example , a person with a family history of heart disease can significantly reduce risk through careful diet , exercise, smoking cessation, and other behaviours.
- The use of drugs to control high blood pressure is also an example of an environmental intervention that reduces the overall risk of heart disease. Heart disease is a typical example of a complex trait influenced by multiple genes as well as many environmental factors.
- Most of the variations found in human beings fall within this category, including personality and other behavioural characteristics. Some traits are more strongly influenced by genetic factors than others, and it is extremely difficult to sort out trait forms that could have a single cause.
- The boxes labelled mild, moderate, and so on represent different severities in which the trait can be expressed; they are analogous to the different forms of anaemia. There are three genes and three environmental factors across the top that influence the trait.
- Heavy lines are major effects, thin lines are minor. If the four types of trait expression were considered to be a single entity without distinction, the genetic and environmental cause could only be characterised as “three genes and three environmental factors, each with major effects.”
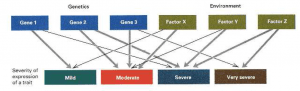
- However, when the different degrees of severity of the trait are considered separately, the situation can be clarified. For example, the mild form is determined by a major genetic factor and a minor environmental factor, and the very severe form is determined by a minor genetic factor and a major environmental factor.
- However, the true complexity remains moderate and severe: the moderate form is determined by two major genes together with one major and one minor environmental factor, and the severe form is determined by two major environmental factors along with one major and one minor genetic factor.
- Figure 4 also illustrates the more general point that features are not already classified in the most informative manner. Progress in genetics has often resulted from proper subdivision of complex traits into distinct types that differ in their genetic or environmental causation.
REFERENCES: